Revised ( R1.1) Content update ( R 1.2 )
Albert’s Lyrebird Menura alberti
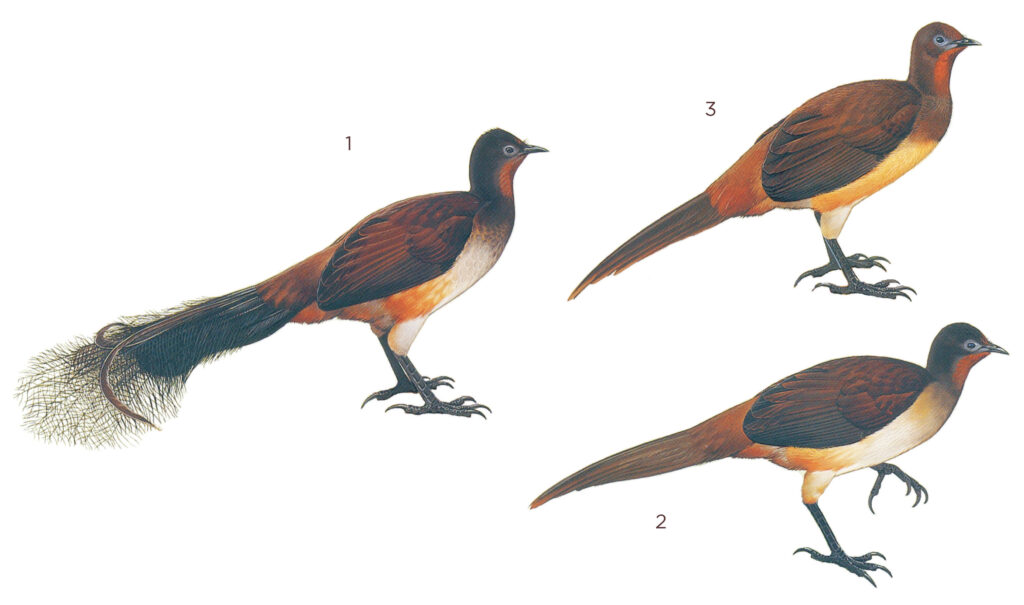
1. Adult male, 2. Adult female, 3. Juvenile
See also Multimedia.
- Status by distribution:
- Global: Least Concern (IUCN)
- Australia: Least Concern (APAB), Not Listed (EPBC)
Menura alberti Bonaparte, 1850, Consp. Gen. Av. 1: 215 — Australia = Turanga (=Terania) Creek, Richmond River, New South Wales.
Latham (1801, Index Orn., Suppl., p. 61), gave the name ‘New Holland Menura’ to the Superb Lyrebird Menura novaehollandiae because of the numerous transparent lunules contained in the broad inner webs of the outer tail-feathers (Greek μήνη, the moon; οὐρά, the tail). The present species was named for Franz August Karl Albrecht Emanuel Prince of Saxe-Coburg and Gotha (1819–61), widely known as Prince Albert, consort to Queen Victoria.
Other English names: Prince Albert Lyrebird, Northern Lyrebird.
Monotypic
. . . . .
Field Identification ⓘ
Length 90 cm (86–93, including tail 47–57 cm long in males, and 36–42 cm long in females); wingspan 77 cm (76–79); weight c. 930 g. Large, mostly ground-dwelling bird. Very similar in size and shape to Superb Lyrebird Menura novaehollandiae but slightly smaller and richer and more extensively rufous plumage, with less elaborate tail in adult male. Presence of extensive tail feathers, including the two lyrates for most of the year comprises the main sexually dimorphic trait; otherwise alike in adult and juvenile plumage, with sub-adult males and females often difficult to visually distinguish. Powerful songs, incorporating much mimicry, distinctive and primarily performed by males though females also possess capability, including for mimicry. Juvenile separable in close view; similar to adult female but with richer rufous-brown chin, throat and foreneck.
Adult male
Head and neck, dark grey, washed red-brown on forehead, and with prominent rufous-brown area on chin, throat and foreneck. Rest of upperbody and folded wing, dark red-brown to rufous-brown, slightly brighter on rump and uppertail-coverts, and grading to duller and greyer on folded primaries. Rufous-brown of foreneck grades into grey-brown on upper breast, in turn contrasting with much paler light grey-brown of lower breast, belly and foreflanks. Rear flanks, vent and undertail-coverts, bright rufous-brown, contrasting strongly with pale belly. Spectacular tail composed of:
- central pair of long, ribbon-like dark-brown median plumes;
- six pairs of long, filmy and luxuriant filamentary feathers, which are black-brown above and dark grey below; and
- long, broad fully webbed outermost pair of lyrates, which are black-brown above and light to dark grey below.
When walking, tail carried in upward-curving train; when bird displaying, tail inverted and spread over body and head, and shimmered. Underwing, dark brownish-grey. Bill, black. Iris, dark brown or black. Broad orbital ring, blue-grey. Legs and feet, brownish grey to dark grey or black.
Adult female
Differs from adult male only in shape of tail, which is shorter, simpler, slightly drooping and appears more pointed when closed, and lacks the full set of filamentaries [though female caught at Goomburra, Main Range National Park had two filamentaries on left side of tail (F. Blackhouse & A. Maisey, 2018)]; tail composed of pair of long, narrow and tapered median plumes, and rest of tail composed of fully webbed, broad brown feathers, with rounded tips.
Juvenile
Similar to adult female; best distinguished by:
- more uniform and richer rufous-brown area on chin, throat and foreneck, and brighter red-brown wash on forehead and forecrown;
- upperbody slightly paler;
- feathers of rump, lower belly and vent have softer downy texture;
- some appear little different from adult in colour and pattern of lower breast, belly and flanks, but others have these areas uniform light rufous-brown and are clearly more extensively rufous below;
- most importantly, tail feathers (excluding central pair of medians) are distinctly narrower, more tapered and pointed.
Immature male
In first year, similar to adult female, from which doubtfully separable in field. With increasing age thought that shape and structure of feathers of tail become progressively more like those of adult male, but plumage changes with age poorly known (see Plumages), and field characters of older immature males undescribed.
Immature female
Poorly known; not easily separable from adult female in field.
Similar species
Likely to be confused only with Superb Lyrebird, the two species not known to occur in the same areas though minor confusion exists in this regard (see Distribution); Albert’s distinguished by:
- brighter and more extensively rufous coloration: mantle to uppertail-coverts, scapulars and folded wing, rufous-brown, contrasting with dark-grey head, neck and uppertail (Superb is predominantly dark grey or grey-brown above, with rufous tones restricted to much duller red-brown on folded wing);
- bright rufous-brown rear flanks, vent and undertail-coverts (dark grey in Superb), last two prominent when tail held over back while male displays;
- chin, throat and foreneck, bright rufous-brown in all plumages (dark grey with, at best, faint red-brown wash in adult Superb, but juvenile and younger immatures aged 1–3 years little different from Albert’s);
- adult male has rather simple uniformly dark-grey outermost (lyrate) rectrices (cf. spectacular rufous-notched, black-tipped and strongly S-curved lyrates diagnostic of adult male Superb).
Voice sometimes also helpful in identification: only Albert’s utters Gronking Song whereas only Superb utters Plik Song (see Voice accounts for details).
Notes
Usually occur singly or in pairs; sometimes in groups of three. Wary and difficult to approach, appearing more so than Superb Lyrebird; more often heard than seen; mostly sighted walking quietly along forest trails; when disturbed, half-run and half-fly to cover or ascend tree in series of leaps. Mainly terrestrial; forage by scratching through leaf-litter and soil in cool moist areas of forest floor. Male sings and displays from platform of trampled vines, or from ground, fallen branches, logs or rocks. For discussion of rather complex vocalizations, see Voice section.
Habitat ⓘ a-z
Mostly confined to rainforests and wet sclerophyll forests with mesic understorey; do not occur in dry sclerophyll forest (Chisholm 1957, 1960; Curtis 1972; Beruldsen 1973; Holmes 1987; Smith et al. 1994; Robinson & Curtis 1996; Schodde 1996; Curtis 1998; Aust. Atlas; A.M. Gilmore). In gullies, along watercourses, and on slopes and ridges of steep mountain ranges; mainly at altitudes >300 m asl though some still recorded to 100 m asl (White 1910; Favaloro 1931; Wigan et al. 1938; Chisholm 1960; Trudgeon 1969; Schodde 1996; A.M. Gilmore; G. Holmes).
Most information from preliminary analysis of results of detailed study of habitat throughout range in NSW (400 sample sites in Tweed, Richmond and Clarence R. Valleys) by A.M. Gilmore. Only found in forests with mesic understorey, either rainforest or wet sclerophyll forest, and then only wettest examples of each. Plant species composition does not appear to be important except that a canopy of eucalypts is always associated with higher densities, compared with rainforest lacking eucalypts (for sites with equivalent climate). Densities increase along a gradient of increasing rainfall and decreasing mean annual temperature; with decreasing moisture index, densities of males decline and individuals increasingly restricted to areas closer to gullies. In comparisons of wet sclerophyll forest and rainforest with equivalent climate and moisture index, higher densities always in wet sclerophyll forest associated with greater weights of litter and logs and slower rates of litter decomposition. Display sites (see Social Behaviour) usually surrounded by dense foliage at ground-level, which typically found below gaps in canopy, which similar to situation in Superb Lyrebird; in optimum habitat, forage and sing up to major ridges whereas in medium quality habitat foraging tends to be confined to lower slopes and gullies. Do not forage or display in dry sclerophyll forest (A.M. Gilmore).
In short-term fauna survey of the Murwillumbah Management Area, ne. NSW (extending from Lismore, along Wilsons R., to Broken Head in the S, N along coast to the Qld border, and W to Tweed Ra.), found mainly within rainforest and wet eucalypt forest with a rainforest substage. Highest densities (in winter surveys) recorded in forest with closed canopy, between 20 and 40 m tall, and with dense understorey 0.5–3 m tall; includes: simple and complex montane rainforest and rainforest thickets; regenerating rainforest under emergent eucalypts, such as Blackbutt Eucalyptus pilularis; montane forest with rainforest understorey; montane acacia forest; mature moist Blackbutt forest; and gully acacia forest (Schodde 1996). See Schodde (1996) for other habitats where infrequently recorded. Note that methods used in this survey may not be appropriate for determining fine habitat for this species.
More general references also indicate preference for rainforest with a dense understorey of vines and shrubs, or wet sclerophyll forest with a dense understorey of rainforest plants(Chisholm 1957, 1960; Curtis 1972; Beruldsen 1973; Holmes 1987; Smith et al. 1994; Robinson & Curtis 1996; Schodde 1996; Aust. Atlas; H.S. Curtis; G. Holmes), including temperate rainforest (G. Holmes). Occasionally found in mixed eucalypt forest, of gullies and lower slopes, with a mesic understorey; and with small amounts of rainforest in wet gullies (Holmes 1987; Reilly 1988; Brouwer & Garnett 1990; Smith et al. 1994).
Distribution and Population ⓘ
Endemic to Aust. Confined to small area of far south-eastern Qld and far north-eastern NSW.
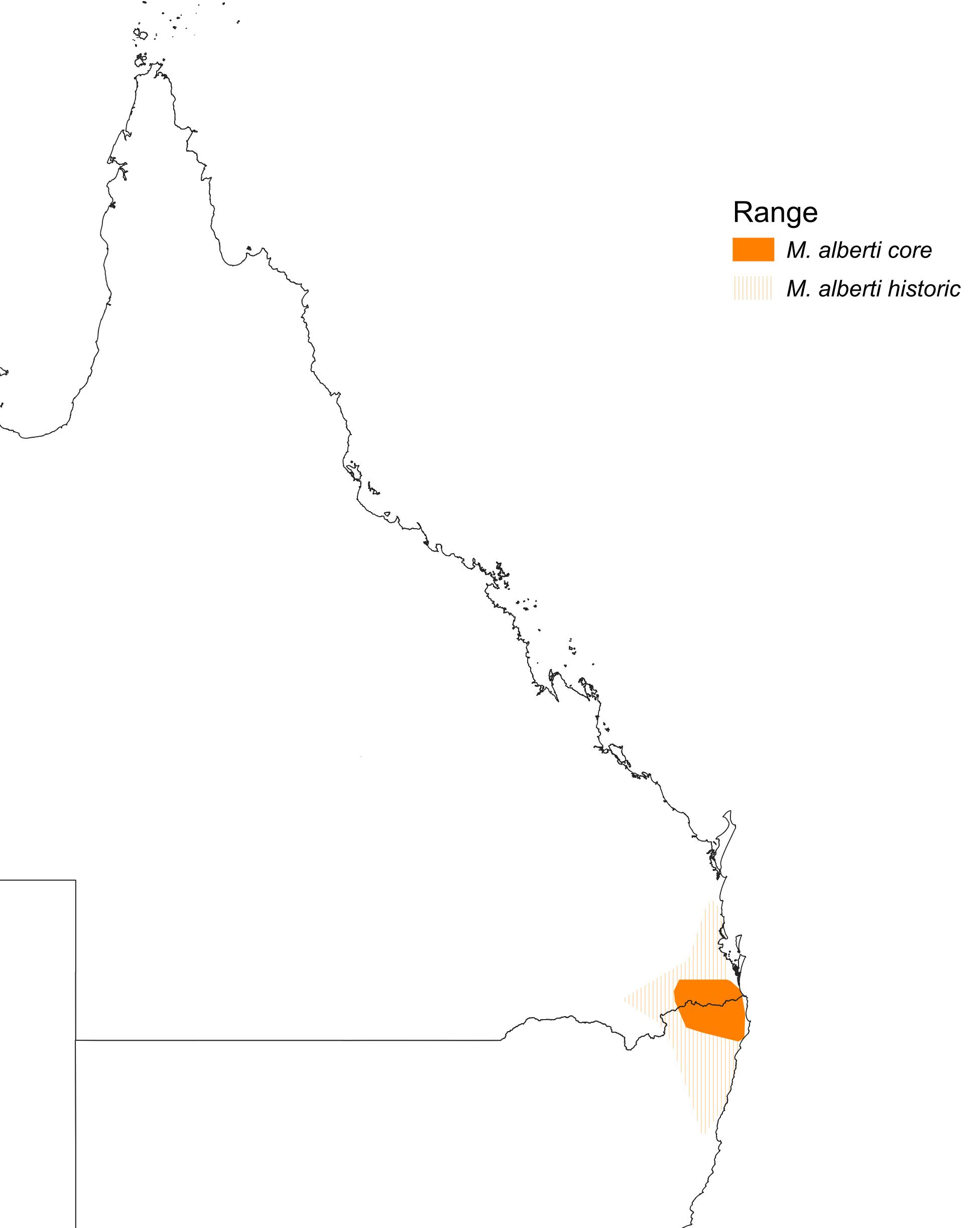
Link to map from book.
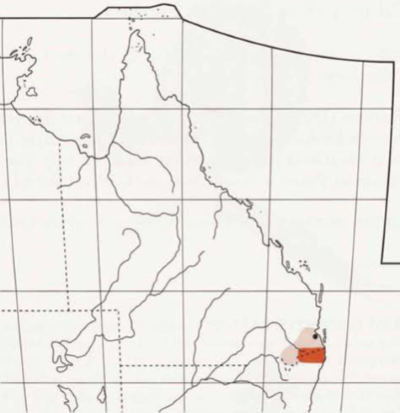{kind=link}
Australia:
- Qld Range restricted to scattered, sometimes isolated, sites in area from Tamborine Mountain to Lamington National Park, and McPherson Ranges. West to Mt Barney National Park and near Killarney. Also found along Great Dividing Range and associated ranges, from Mount Superbus and Cunninghams Gap, in Main Range. National Park, North to the Mistake Mountains (Curtis 1972; Beruldsen 1973; Wheeler 1973; Roberts 1979; Robinson & Curtis 1996; Storr 19; Aust. Atlas; L. Nielsen). Reports from Blackall Range (Nielsen 1959b; Chisholm 1960) almost certainly refer to misidentified calls of Satin Bowerbirds (L. Nielsen).
- NSW Confined to Northern Rivers Region. Mostly along McPherson and Tweed Ranges, from Wollumbin National Park, West to Border Ranges National Park, and as far West as at Acacia Gap in Koreelah State Forest (just East of Legume), and extending South to Koonyum and Nightcap Ranges in East (near Mullumbimby, including Whian Whian State Forest and Nightcap National Park), West to Boundary Creek, South East of Kyogle, and Toonumbar State Forest in Richmond Range (Trudgeon 1973; Morris et al. 1981; Robinson & Curtis 1996; Schodde 1996; Aust. Atlas; NSW Bird Reps; G. Holmes). Isolated population farther South, at Uralba Nature Reserve, in Blackwall Range, between Alstonville and Bagotville (Gosper 1986; Holmes 1987; G. Holmes). Report much farther West, at site North East of Tenterfield, 16 Oct. 1992 (NSW Bird Rep. 1992), considered highly doubtful (H.S. Curtis; A.M. Gilmore) and suggested probably misidentified Superb Lyrebird (H.S. Curtis).
Breeding
Throughout range.
Change in range, populations
Populations at Tamborine Mountain said to have declined greatly (Mack 1952; Roberts 1977; L. Nielsen); also thought to be declining in some other areas (Trudgeon 1973; Roberts 1979). Numbers ‘a few miles from Brisbane’ said to have declined by late 1920s (Roberts 1930). Formerly more widespread at lower altitudes at turn of 20th century (Holmes 1987; Schodde 1996). Said, probably fancifully, to have once occurred North to Wide Bay, Qld (Chisholm 1957; Macdonald 1969; Campbell). The historical recordings that exist in relation to this purported northern population include a reported visual sighting from the Blackall Ranges (Chisolm), a name in the indigenous language of the region (Kabi Kabi) recorded as referring to this species (Meston) and a single museum skin with the provenance listed as ‘Mary River’. In lieu of no other evidence to the birds’ historic or ongoing existence in habitat north of the Mistake Mountains, clerical error seems likely in these cases (reference/pers comms).
Status
Of special concern (Garnett 1993).
Populations
No estimates of abundance.
Conservation Status ⓘ
Albert’s Lyrebird Menura alberti
APAB 2020 conservation status: Least Concern
Justification of status: the habitat of Albert’s Lyrebird is nearly all managed for conservation and, while fires affected some of its range in 2019, losses are unlikely to be continuing. (Stewart et al. 2021)
Global IUCN Red List status of species: Least Concern
EPBC Act status: not listed
Legal status in range states: NSW: VU, Qld: NT
Status certainty: medium; high-quality mapping demonstrates how much habitat was burnt severely in 2019 but no dedicated monitoring.
2010 Action Plan status: Near Threatened: change in status in 2020 genuine: earlier sustained declines due to forestry in what is now Whian Whian Conservation Area have ceased; non-genuine (guidelines): IUCN Red List definition of location tightened.
Threats and Human Interactions ⓘ
Thought to have been adversely affected by clearance of rainforest for agriculture (Irby 1930; Chisholm 1957; Smith 1968; Roberts 1977; Garnett 1993; Curtis 1998). Also possibly adversely affected by clear-felling and fragmentation of rainforest habitat below 300 m asl (Schodde 1996); and also possibly by harvesting of timber from rainforests (Nielsen 1959a). However, in Murwillumbah Management Area (see Habitat), habitat requirements possibly met in moderately mature rainforest and wet sclerophyll forest at higher altitudes, under a selective logging regime with 20–50 years between logging treatments and which excludes fire (Schodde 1996). In north east NSW, densities can be as high in some regrowth forest as in old-growth forest, but in forests susceptible to dominance by sedges and ground-cover plants, or to invasion by Lantana after logging, densities are lower than expected. Further, densities are lowest, often zero, in plantations of eucalypts or Hoop Pines established in areas that were optimal habitat. Hoop Pine plantations now occupy large areas of the Nightcap Range National Park, reflecting its transition from State Forest in the 21st century. These areas, though now protected, presumably offer limited potential habitat for the species. Thus, trend of increasing intensity of forest management likely to cause further declines (A.M. Gilmore). Much of the species’ habitat is surrounded by very dense rural settlement, and densities of the species is lower than expected in rather large areas of potential habitat in forest margins adjoining such settlement (A.M. Gilmore). Much of the species remaining habitat reserved.
Formerly shot, either to be supposedly eaten in pies (Trudgeon 1973), to supply tail-feathers to ‘globe-trotting curio-hunters’ (Jackson 1909), or by vandals (Marshall 1935). Reports of frequent human consumption should perhaps be treated sceptically as mentions are scarce and the bird’s longevity would likely not render their flesh particularly desirable fare. Habitat destruction or alteration likely the major cause of diminished numbers. In north east NSW, vandals once said to have surrounded patch of scrub and set it alight, shooting Lyrebirds that tried to escape; their tails were cut off and the bodies left to rot (Jackson 1909). Thought to be killed and eaten by Foxes and Cats (Trudgeon 1973), though this also thought doubtful (H.S. Curtis). Lyrebirds annoyed people who were erecting the rabbit-proof fence in Qld by filling in trenches which had been dug by workmen the previous day (Vidler 1933).
Movements ⓘ
Sedentary, with males and females remaining in same general area year round (Schodde 1996; Aust. Atlas; A.M. Gilmore; see Social Organization); though also described as resident in NSW (Morris et al. 1981). While poorly known and no studies, some dispersion or local movement beyond territorial boundaries may occur outside breeding season, by adult males and females and fledged young (Aust. Atlas; H.S. Curtis). However, claim that forage more widely and in different habitats in winter, and withdraw to montane rainforest areas in summer (Schodde 1996), considered doubtful because many territories do not abut montane habitat, and may be up to 40 km from such habitat (A.M. Gilmore); and birds breed in winter, when they are restricted to territories or nest-sites (see Social Organization, and Breeding: Site). Crossed 180 m of open forest and 140 m of potato cultivation to reach isolated patch of rainforest on Acacia Plateau, north east NSW (Trudgeon 1973).
Banding
No verified information on banding. Supposedly of five banded in Aust., 1953–97, none recovered.
Longevity: Longest lived estimated to be a male holding territory in Lamington National Park, Qld, for at least 15 years, 1984–99, and already adult when first observed; suggested that probably ≥22 years old (H.S. Curtis). This bird, dubbed ‘George’, lived at least another decade, suggesting an age in excess of 30 years.
Food ⓘ
Very poorly known. Diet and foraging behaviour probably similar to that of Superb Lyrebird, but no details. Appear to eat mainly insects and their larvae, and other soil-dwelling invertebrates. Camera footage from Carabeen Nature Reserve has also shown feeding on earthworms, though this is apparently rare; one record of female taking earthworms that were placed out for her, but she fed them to nestling (Lindsey 1994). Mostly forage singly (Keast 1944). Feed on ground, preferring areas with deep moist leaf-litter and fallen logs (Jackson 1920, 1921; Keast 1944; Chisholm 1957; Lindsey 1994; Schodde 1996; H.S. Curtis; A.M. Gilmore). In study throughout range in NSW (see Habitat), typically in areas that are rather open and lack dense shrub cover but have well-developed taller strata (A.M. Gilmore); these observations supported by other, more general, references (Jackson 1920, 1921; Keast 1944; Chisholm 1957; Lindsey 1994; Schodde 1996; H.S. Curtis). Scratch among debris, turning over leaves for invertebrate prey (Jackson 1920, 1921; Keast 1944; Chisholm 1957; Lindsey 1994; Schodde 1996; H.S. Curtis); also observed digging down into soil, up to 5 cm below surface (H.S. Curtis). Gould provides only positive record of feeding, on small insects, mainly beetles, obtained from ground by scratching and digging on forest floor. Chisholm (1957) quoted observations claiming snails as part of diet, but Schodde (1996) suggests that soft pliable bill of Lyrebird may not cope well with large gastropods. Also claimed that birds seek cockroaches, worms, land snails, and other invertebrates; but were not actually seen capturing or eating any of them (Lindsey 1994); and also that they eat insects, worms and land snails (Aust. RD).
Social Organisation ⓘ
Little known and no detailed studies, though some detailed information from various long-term observations (Curtis 1972, 1998; Robinson & Curtis 1996; Schodde 1996; H.S. Curtis; A.M. Gilmore; L Nielsen). The vast majority of what is known about this species relates to the male’s vocal displays during the courtship process, reaching its peak in winter.
Video recording of some aspects in Behrens (1994) with further footage of female’s raising of chick from Lamington National Park (Threlfo). Much footage gathered from Carabeen Nature Reserve through use of trail cameras (Fearnley). Little known in Albert’s lyrebird compared with Superb Lyrebird, particularly with the species’ propensity to only display in dense vegetation cover, and general and secondary sources often extrapolate from Superb to this species, without solid evidence that this warranted; we have been cautious in use of such sources. Sources of much information in Schodde (1996) not always certain. Very difficult to observe, with most sightings merely glimpses; in contrast, vocalizations powerful and well known, and birds much more often heard than seen (H.S. Curtis; see Voice).
Solitary. In breeding season, solitary, except when male and female copulate or during territorial confrontations. Outside breeding season, males may tolerate one another’s presence, and, very rarely, 2–3 seen together (H.S. Curtis). Other reports usually of singles (Keast 1944; Gould; H.S. Curtis; L. Nielsen). Rarely, two female or immature-type birds seen together, at least sometimes being female and her offspring (H.S. Curtis; L. Nielsen); and, even more rarely, groups of three observed (Jackson 1921; Favaloro 1931; Clarke 1968).
Bonds
Beyond some rudimentary knowledge of the female’s rearing of the chick, largely unknown and much of what is written is speculative, with little or no supporting data. Said that single males sometimes associate with single females, both during and outside breeding season, but also suggested mating system probably polygynous, where territorial males mate with itinerant females from adjacent territories (Schodde 1996). In north east NSW, mapping of calling males during breeding season, indicated that males had an over-dispersed spatial pattern much as other territorial passerines, and no evidence of lekking behaviour (A.M. Gilmore). However, has been claimed that males maintain territories in dispersed leks, in which they sing and display (Schodde 1996; Robinson & Curtis 1996) with leks often on, or near, high ground, such as tops of ridges (H.S. Curtis), but no studies or supporting evidence available and further study needed. Territories of males at least sometimes clumped within available habitat (Reville 1973; H.S. Curtis; D.N. Jones). Males in an area can call at the same time and are in auditory contact with one another (Reville 1973; Schodde 1996; H.S. Curtis; D.N. Jones). Also thought that might be promiscuous, as in Superb Lyrebird (q.v.) (Schodde 1996). Before laying, female visits male to mate and then returns to her nesting area (L. Nielsen). Age of first breeding not known (H.S. Curtis). Male thought to take no part in parental care (Lindsey 1994; Schodde 1996; Curtis 1998; see Breeding).
Breeding dispersion
Closest active nests found in one study in south-eastern Qld were c. 80 m apart (L. Nielsen). Partly built nest-platforms sometimes found, often in newly occupied territories, and may be made by young non-breeding females (Beruldsen 1973); but also sometimes found in areas known to have been occupied by same female for some time (L. Nielsen).
Territories
Males territorial when breeding (Robinson & Curtis 1996; Schodde 1996; A.M. Gilmore) and seem to spend most of their time in their territory (L. Nielsen). Within territory, male displays from platforms of vines and twigs and does not build mounds (see Social Behaviour). One mature male, apparently the same bird, occupied same area at Lamington National Park each breeding season from c. 1984 and was still there in 1999 (H.S. Curtis); this same territory seems to have been occupied by a male 70 years earlier (Ratcliffe 1938; H.S. Curtis). Little is known of dispersion of females, and whether they are territorial or not, but it is likely that they are; also likely that female and chick leave nest-area once chick is old enough (H.S. Curtis). Males and females probably maintain separate territories in breeding season (L. Nielsen). At least some females return to the same nesting areas each year (L. Nielsen). Relationship between areas where females nest and territories of males not known (L. Nielsen contra Strahan). Birds can leave territories and wander locally after breeding season (Aust. Atlas; H.S. Curtis). Breeding territories break down from late spring to early or mid-autumn when breeding adults, non-territorial adults and immatures forage throughout local areas, though breeding males and females keep to same general areas year round (Schodde 1996).
Size of territories (all males): At Lamington National Park, three territorial males in c. 25 ha of suitable habitat; at Toonumbar State Forest, all display platforms of one male were within 6 ha but, as platforms are usually towards the middle of the territory, territory was probably 8–10 ha; at another site in an isolated patch of forest, eight calling males in c. 12 ha, in July, though birds may have been crowded because patch isolated (H.S. Curtis); territories said usually to be 5–15 ha (Schodde 1996). In detailed study in north east NSW, little variation in size of home-ranges, though density can vary with habitat (A.M. Gilmore); cf. claim that size of territories or home-range probably varies with productivity of habitats (Schodde 1996). Along a track in Mistake Mountains, a singing male heard every 100 m (H.S. Curtis); at another site, up to 12 males heard calling at one time during breeding season (Nielsen 1960); at another, three males heard calling nearby, and two others in the distance, in July (Beruldsen 1973); at McPherson Ranges., four singing males 400–750 m apart in June (G. Holmes); at Lamington National Park, in October., different birds heard calling every 800 m or so (Robertson & McGill 1948); across multiple locations, displaying males found on average 300 m apart (F. Backhouse); said that calling birds seldom closer than c. 400 m (Gould). Said that males stay at least c. 370 m apart (Smith 1988).
Roosting
At night; in trees (Keast 1944; Schodde 1996; Curtis 1998). In breeding season, territorial males roosts alone within territory (H.S. Curtis), and males seen to go to and leave roost alone; females roost in nest with young, even when young are quite large; after young fledge, female and young possibly roost together (L. Nielsen). No evidence that ever roost as breeding pair (H.S. Curtis; L. Nielsen) or in groups (L. Nielsen), contra claims that commonly roost in small dispersed groups of several birds or, in case of breeding adults, usually as pair (Schodde 1996); basis of latter statements not known. Roost on sheltered branches within subcanopy of mature forest, 7–20 m above ground (Schodde 1996); territorial males usually roost in canopy, often >20 m from ground (H.S. Curtis); twice recorded roosting at top of Turpentine Syncarpia glomulifera in subcanopy of Blackbutt forest (A.M. Gilmore); once c. 10 m up a tree (Keast 1944). In morning in breeding season, mature males call from roost, staying there or in the canopy until well after dawn (H.S. Curtis); during hour before dawn, calling virtually continuous, except when birds moved between calling sites (A.M. Gilmore). In June, one male began to sing from roost at 06:08, then flew to ground at 06:15 (Robinson & Curtis 1996); another male began to sing at 06:16 and went to ground at 06:24 (H.S. Curtis). Males seen leaving roost-trees at first light, and ascending them at last light (L. Nielsen). Perch on stumps, logs, low hanging vines and in trees during day (Marshall 1935; Robertson 1948; Trudgeon 1969; Beruldsen 1973; Robinson 1974).
Social Behaviour ⓘ
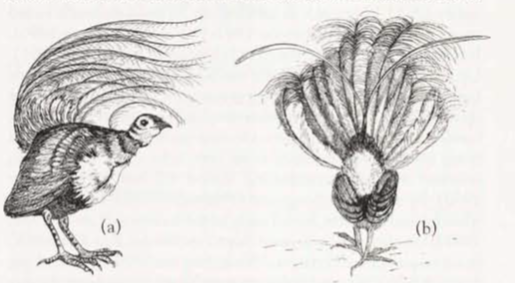
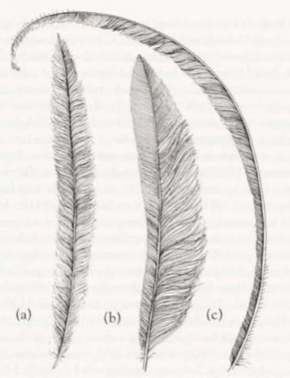
Some published information but not well known; sources as Social Organization. Extended observation difficult as birds usually extremely shy and, unless approached very cautiously, birds usually leave silently (e.g. Jackson 1920; Marshall 1935; Keast 1944; Thomson 1964; Curtis 1972; Lawler 1980; Smith 1988; Robinson & Curtis 1996; Curtis 1998; Gould); said to be less curious than Superb Lyrebird (Favaloro 1931). However, can occasionally become tame (Curtis 1998). Behaviour, including nesting behaviour, appears similar to that of Superb Lyrebird, but male Albert Lyrebird usually displays on an often inconspicuous display platform, and not on a mound (Curtis 1972; Smith 1968, 1988; Gould). Long tail also not so elaborate as that of Superb (see Field Identification, Plumages). In first phase of display on platform, tail held more erect (Reilly 1988); in second phase of display, when tail spread and held over body and head, resulting fan is dome-shaped, and differs from that of Superb because curvature of the feathers of each species differs; see Figure 1 (Curtis 1972). Main feature of display is singing and dancing by individual males on display platforms within territory (Schodde 1996).
Display platforms and clearings: Each male usually displays while standing on one of a number of display platforms or clearings that he uses regularly (Curtis 1972; Schodde 1996). Display platforms are usually close together; and male feeds and roosts near them (L. Nielsen); males known to use the same platforms each year (H.S. Curtis; L. Nielsen). Display platforms typically made of several thin vines or fallen branches lying on or just above ground and crossing each other; platforms well concealed and surrounded by screening vegetation from above, so that display not often seen by people. Where available, often in Lawyer Vine Calamus muelleri, which provides vines for both platform and surrounding screen; if prickly Lawyer Vines used in platform, prickles are worn off. In north east NSW, also often display in Lawyer Vine and in clumps of Native Ginger Alpinea caerulea (A.M. Gilmore). In vicinity of Killarney in Qld, Native Grape (Cissus antarctica) is preferred vine species in the absence of Lawyer Vine. On Koonyum Range, clumps of bogrush (Schoenus sp.) used as platforms (F. Backhouse).
Ground below platforms often scratched clean, and where male stands on platform is often marked with mud (Curtis 1972). Male might manipulate vegetation and vines to form platform (Curtis 1972; Lawler 1980). Display platforms provide a clear and slightly elevated area for display (Robinson & Curtis 1996). Birds sometimes call and display elsewhere, such as on rocks, logs, fallen branches or bare ground (Curtis 1972). Also seen displaying in: open space in dense undergrowth (Favaloro 1931); small circular clearing among ferns with little screening vegetation (Beruldsen 1973); simple mound in rather open area with small pile of mud-stained twigs scratched from floor of forest (Lawler 1980); in scrapes that bird has cleared for up to 1 m round (Schodde 1996); notes in Gould of birds displaying in holes in ground have not been reported since and seem most unlikely. Clearings round display areas possibly result of constant use rather than deliberate clearing by bird (L. Nielsen). Display sites are surrounded by dense foliage near ground; such dense foliage typically occurring where there are openings in the forest canopy, and is similar to situation in Superb Lyrebird (q.v.) (A.M. Gilmore).
Display sites occur within rainforest or in tall wet sclerophyll forest close to ecotone between rainforest and sclerophyll forest (Schodde 1996); also occur on ridges in wet sclerophyll forest where mesic understorey is intact and not altered by forestry or fire management practices; do not display in dry sclerophyll forest (A.M. Gilmore). Vocal displays, in particular loud Territorial Song, used often by adult males, and probably, at least in part, adaptation to territorial defence and attraction of mates in dense understorey of forest. Calls indicate territorial status, onset of copulation or presence of predator or disturbance; Territorial Song may provide communication with distant males, whereas Sequential Song may be targeted at nearby birds, male or female (though it is regularly interrupted by Territorial Song). During breeding season, males spend up to 4 h/day singing, with sessions lasting from a few minutes to >1 h; many, but not all vocal displays accompanied by visual displays (Curtis 1972; Robinson 1991; Robinson & Curtis 1996; see Voice). After start of breeding season in late Apr.–early May, display and copulation said to intensify through May to mid-June with diminished singing by males late June–early Aug, during nest-building and incubation (Schodde 1996). Sequential Song said to be culturally transmitted, see Voice. At rest, tail curves upward (Curtis 1972).
Agonistic behaviour
Territorial advertising
Male gives very loud Territorial Song (also known as ‘Whistle Song’: Backhouse et al 2021), possiblyto communicate with distant, neighbouring males, and is usually answered by neighbouring males. Territorial Song carries far, up to 500 m through vegetation (see Voice). Before descending trees in early morning, male may give variations of Territorial Song; once on ground, often moves close to boundary of territory and calling includes much of his Territorial Song at first; if there is no intrusion by another male, he moves to a more central part of territory and there mainly gives Sequential Song with infrequent Territorial Song (Reilly 1988). Birds with territories on large slopes tend to move from roosting sites in lower parts of territories to progressively higher calling sites (A.M. Gilmore). For details of frequency of singing, see Voice (Song). During breeding season, taped playback of either Territorial Song, Sequential Song or Gronking provoked territorial defence in males: bird usually stopped singing, approached sound, and went to display on nearby display platform, giving full Sequential Song, stopping now and then to listen or give Territorial Song. Rarely, when recordings made by leaving a microphone at display platforms, there were sounds consistent with another Lyrebird being present and a dispute taking place; calls were often harsh and apparently threatening; normally, vocal displays contain none of these sounds, though alarm calls do occur in most displays of Superb Lyrebirds (Robinson & Curtis 1996).
Threat
One encounter in Nov., between an adult male and a smaller bird, possibly also male, was probably aggressive and involved raking ground with claws; birds also faced each other, with tails spread over body (see below), wings spread out and lowered (at least in larger bird) and necks stretched out and in line with bodies; both then stepped exaggeratedly till bills almost touched; at this point, larger bird gave Gronking Call, jerking wings down each time he called. Birds also chased each other, with larger male holding tail erect; larger male also gave a quilp call while displaying tail with legs bent and body bent forward (Clarke 1968). Others have seen birds chasing each other (Favaloro 1931) though circumstances not known.
Alarm
When disturbed, may give one of a variety of calls (see Voice), move to a nearby tree and hop from branch to branch, up to c. 15 m high or to top of tree, and watch or glide or move away; may just run away (Jackson 1921; Robertson & McGill 1948; Curtis 1998; Gould); may just call and quickly leave (Robinson & Curtis 1996). Often, if a person is nearby and is seen, a male will quickly move away, but if person approaches carefully and is only heard, bird does not move (H.S. Curtis).
Sexual behaviour
Vocal displays directed toward females are quieter than those directed toward males; most Sequential Song is directed to nearby females, though it often includes mimicry of loud Satin Bowerbird Ptilonorhynchus violaceus advertising call which may be directed to distant females (Robinson & Curtis 1996).
Display with tail: (Fig. 1) In first phase, bird suddenly raises and partly fans tail; outer tail-feathers (lyrates) are spread apart so that bases of feathers at angle of c. 120° and rest of tail spread within shafts of outermost pair of filamentiares, which are at angle of c. 80°; basal half of tail held almost vertically, but natural curve of tail results in distal half lying over bird’s back; posture shows bright rufous patch at base of undertail, contrasting with shimmering silver undersurface of rectrices; bird sways body from side to side and moves head continually, allowing it to see surrounding area, but feet move little. Much of the display is given with tail in this position (Curtis 1972; H.S. Curtis). In second phase, bird throws tail farther forward, and fans it 180°, so that silver dome of feathers covers head and body (but bird still able to see well); outside rectrices are horizontal at bases but distals ends curve forward and down below body; bird gives high steps from foot to foot and occasionally whole tail shimmers (Curtis 1972; Lawler 1980; Reilly 1988). Bird sings throughout display (H.S. Curtis). Displaying with tail, dancing and singing continues for 20–30 min (Beruldsen 1973; Lawler 1980). When performing on platform, stepping on the slightly raised vines of platform causes them to move, which disturbs adjacent vegetation and sometimes shakes foliage several metres away (Favaloro 1931; Curtis 1972; Lawler 1980); bird steps from one foot to the other or moves one foot p and down, alternately depressing and releasing vines, causing movement of vegetation (Reilly 1988; Backhouse et al 2024); video of display shows bird grasping vine and moving it up and down to produce movement (Behrens 1994); during Rhythmic Gronking Song, movement of vegetation is regular and synchronized with rhythm of the song (Robinson & Curtis 1996; Backhouse et al. 2024). Displaying males probably supplement their signalling by manipulating vines (Robinson & Curtis 1996). Gould noted that bird lowers wings when displaying with its tail. Sequential Song seems directed at females: after one male left roost, and female known to be in area, he sang for 33, 4 and 65 min respectively at three different display platforms, giving 32, five and 84 Sequential Songs and only eight, one and seven Territorial Songs; he ignored neighbour’s Territorial Songs, which normally would have stimulated response; performance ended with Clicking and copulation thought to have occurred (Robinson & Curtis 1996; Curtis 1998).
Copulation
Said that one male sang and displayed till female presented herself; he then began slow clucking noise which became faster as copulation proceeded; afterwards, birds left together, male clucking softly alongside female; within a few minutes, male began territorial calling again. Occurred in Mar., outside normal breeding season (Smith 1988). One male recorded during copulation observed singing Gronking Song in the lead-up to copulation, then mimicry of Grey Shrike-thrush Colluricincla harmonica and Mountain Brushtail Possum Trichosurus cunninghami during copulation. Copulation was followed by ‘Clicking’ sounds, similar to those found in Superb Lyrebirds (F. Backhouse, recorded 11 July 2018).
Relations within family groups
Male appear to take no part in parental care of nestlings or fledgelings (Lindsey 1994; Schodde 1996; Curtis 1998; see Breeding).
Parental anti-predator strategies
Often nest on steep slopes where they can glide away from nest if predator approaches (H.S. Curtis). Typically, when brooding female disturbed on nest, she gives a single loud scream as she flies from nest (Beruldsen 1973; L. Nielsen). In response to people, females often disappeared when nestlings approached; one female perched near nest, partly hidden by leaves, and made intermittent alarm calls; when nest approached more closely, she would fly down to distract observers and give a different alarm call, to which chick responded with a loud screech; a male approached, and seemed to make soft calls, but the significance of this is not known, and may have just been coincidental (H.S. Curtis). At another nest, female flushed and when nestling more closely approached, young squealed and jumped from nest; female approached observer closely and gave guttural calls that included mimicry (G. Holmes). When with nestlings, some females become much more tolerant of people (L. Nielsen).
Voice ⓘ
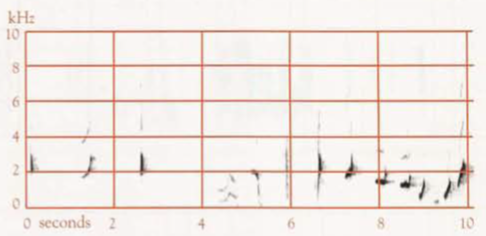
The most well-known behavioural trait. Detailed studies of vocalizations of male, including sonagrams, by Robinson & Curtis (1996); of regional variation in Territorial Song, including sonagrams, by Reville (1973) and Backhouse et al (2021); of musical values of Song by Hartshorne (1953); and of variation in mimetic repertoires by Backhouse et al (2022, 2023). Unless indicated otherwise, discussion is of voice of adult male, and statements are from Robinson & Curtis (1996) and contribution by H.S. Curtis. Much more often heard than seen (Keast 1944; Robertson & McGill 1948; Mack 1952).
The Song is most striking, and consists of prolonged Sequential Song (or Recital Mimicry: Backhouse et al 20222, 2023), which is mimicry of vocalizations of bird species and other natural sounds, usually delivered in a rigid order, loud Territorial Song (or Whistle Song: Backhouse et al 2021) and Gronking Song (Ratcliffe 1938; Robinson & Curtis 1996). Territorial Song audible no farther than 0.5 km through rainforest or wet sclerophyll forest, but audible to over 3 km where there is no intervening cover, such as across a deep valley (A.M. Gilmore); at Lamington National Park, said that calls heard up to 1.5 km away (Robinson 1991; Robinson & Curtis 1996). Up to 12 birds uttering Song have been heard from the one location (Nielsen 1960).
Song given most in June and July, for up to 4 h/day, mostly in the morning and evening (Curtis 1972; Lawler 1980, 1981; Gould). On one such day, a bird uttered Song almost continuously from first light until 11:00, then resumed at 13:00, continuing almost till last light (Lawler 1980). By mid-Aug., Song said to have diminished to a series of whistles and crackles, heard early in the morning (Lawler 1980, 1981); on three days in mid-Sept., in Border Ranges National Park, NSW, many sequences of Territorial Song, Sequential Song, Gronking Song, Alarm Calls and Subsong were heard throughout the day (V. Powys). There is much less Song in a dry winter, and it may stop altogether during droughts (H.S. Curtis). Female mainly silent, but she too has a powerful voice, and is capable of mimicry and Song (Curtis 1998).
Geographical variation: Dialects occur in Territorial Song, Sequential Song (including sounds mimicked and temporal order of mimicry and Gronking Song (see below). Song is similar to that of Superb Lyrebird: within Song, the Territorial Songs of the two species can be distinguished with experience, but it is easier to listen for Gronking Songs within the Song, which are made only by Albert’s and, though subject to considerable regional variation, are easily recognized (H.S. Curtis). Respond to playback of own or neighbours’ Song, even to Song from which all species-specific calls have been removed, i.e. consisting entirely of mimicry (Robinson 1974, 1975; Robinson & Curtis 1996). Reaction does not differ significantly between playback of local Territorial Songs and those of other populations (Reville 1973).
Mimicry
Mimicry forms 70% of Songs, with the species’ own calls comprising the rest. Relative frequency of components, the rate and order of delivery, and length of bout are dealt with below under Song. Prefer to mimic brief, often unmusical songs, or mere call notes, or fragments, torn out of their musical context, from the more elaborate and highly organized songs of other species (Hartshorne 1973). For all areas studied, several mimicked songs and calls of Satin Bowerbird form by far the largest component of Sequential Song. The Bowerbird’s advertising call is usually included, with local Bowerbird dialects accurately mimicked at Lamington National Park and the Knell Section of Tamboring Mountain National Park, Springbrook National Park, and Main Range National Park, Qld (Putland et al 2006). Unlike most of the Sequential Song, mimicry of these advertising calls is given as loudly as Territorial Song, and a common experience when approaching a singing male in the early morning is to hear at first only Territorial Song and mimicked Bowerbird advertising calls, and then, when closer, to hear the rest of the Sequential Song linking them. A distinct call of the Satin Bowerbird, described as three or four, is mimicked (Clarke & Clarke 1972; H.S. Curtis).
Other species mimicked include: Brown Cuckoo-Dove Macropygia phasianella, Wonga Pigeon Leucosarcia melanoleuca, lorikeets Trichoglossus or Glossopsitta, Australian King-Parrot Alisterus scapularis, Crimson Rosella Platycercus elegans, Laughing Kookaburra Dacelo novaeguineae, Lewin’s Honeyeater Meliphaga lewinii, Yellow-throated Scrubwren Sericornis citreogularis, White-browed Scrubwren Sericornis frontalis, Eastern Yellow Robin Eopsaltria australis, Australian Logrunner Orthonyx temminckii, Eastern Whipbird Psophodes olivaceus, Grey Shrike-thrush Colluricincla harmonica, Paradise Riflebird Lophorina paradisea, and Green Catbird Ailuroedus crassirostris (Chisholm 1946; Nielsen 1959a, 1960; Clarke 1968; Robinson & Curtis 1996; Backhouse et al 2023; V. Powys).
Other sounds mimicked include: wingbeats, rustling of feathers, the thump of feet of a large bird landing on a branch, the snapping of the bill of the Laughing Kookaburra, and calls of frogs (Marshall 1935; Chisholm 1946; Smith 1988; Robinson & Curtis 1996). The number of species mimicked is generally fewer than for Superb Lyrebird (Robinson 1975, 1991). In some areas, introductory notes to the Territorial Song appear to mimic Australian King-Parrots, Eastern Yellow Robins. Grey Goshawks Accipiter novaehollandiae, or, less often, Crimson Rosellas. In Toonumbar State Forest, NSW, certain calls of Pied Currawongs Strepera graculina and Satin Bowerbirds (but not the advertising call) are sometimes used as part of Territorial Songs. Calls of Grey Shrike-thrush Colluricincla harmonica, Eastern Whipbird Psophodes olivaceus, and brushtail possums Trichosurus are mimicked immediately after Gronking Song; and calls of the Eastern Whipbird (not well-known antiphonal call) may be given with what appear to be Alarm and Threat Calls; see below.
Spontaneous mimicry reported only once: a male Satin Bowerbird gave his advertising call and an Albert’s Lyrebird interrupted his Sequential Song (a Satin Bowerbird advertising call was not the next call in the sequence), gave a single perfect imitation of Bowerbird’s call, and then continued the Sequential Song (Curtis 1998). In the breeding season, almost all mimicry occurs within Sequential Song; outside breeding season, a much wider variety of sounds may be mimicked, including birds, mammals, and frogs; the Brown Cuckoo-Dove, Wonga Pigeon, and lorikeets were mimicked only outside the breeding season (H.S. Curtis). There is no reliable report of mimicry of man-made sounds or introduced species in breeding season (Robinson & Curtis 1996; L. Nielsen). Mimicry of crowing of rooster, cackling of hens, barking and howling of dogs, and the noise of sharpening a saw has been reported (Vidler 1933; Gould), but the time of year not given. The Gronking Song heard at a distance could be mistaken for the barking of a dog (Clarke 1968; Reilly 1988).
There is a reliable report, from outside the breeding season, of imitation of the sound of a tractor starting (Chisholm 1965; Curtis 1998). In an area frequented by people, outside breeding season, one bird did a very good imitation of people talking, although, with the exception of ‘Hey Bill!’, individual words were not recognizable (H.S. Curtis). Thought that most mimicked calls culturally transmitted from adult males to young birds (Reilly 1988; Smith 1988; Robinson 1991; Robinson & Curtis 1996). The Australian Logrunner is mimicked in the Blackwall Range, NSW, despite being locally extinct (G. Holmes). Also, Laughing Kookaburra mimicked deep within rainforest, 4–5 km from nearest Kookaburra population (L. Nielsen).
Non-vocal sounds: When on display platform, bird grasps one of the vines or sticks and moves it up and down in time with Gronking Song. If the vine or stick being moved crosses another, and both are hard and dry, the effect of rhythm sticks is produced, the tapping sound of which accompanies the Song. See Social Behaviour for some further details.
Adult male
- Song: Song of males has three main components: Territorial Song, Sequential Song and Gronking Song. Clicking, probably similar in purpose to that of the Superb Lyrebird, is rarely recorded but occurs after copulation (F. Backhouse). Unlike Superb Lyrebird, where Low Level Alarm Calls occur in Song, the Song of Albert’s appears to contain no alarm calls. Much, but not all Song, is accompanied by visual display. Bouts of Song may last from a few minutes to >1 h; sessions lasting 30–50 min are common (Curtis 1972). In early June, at the Knoll Section of the Tamborine Mountain National Park, Qld, one bird began singing from roost, starting at 06:08; at 06:15 he flew to the ground and, at first, gave only Territorial Songs, then some fragments of Sequential Song between Territorial Songs; by 06:25 he had given 30 Territorial Songs and full Sequential Song began; the bird sang for 33 min (32 cycles of Sequential Song and eight Territorial Songs) at the first Display Platform, 4 min (5 and 1) at a second, and 65 min (84 and 7) at a third. At the end of the last full cycle of Sequential Song there was 30 s of Song derived from the Sequential Song but not in the stereotyped order, then 9 s of Clicking, 30 s more of mimicry, and finally 20 s of Clicking. This performance was unusual for the inclusion of Clicking and for the low number of Territorial Songs. At the same time of year at Mt Mistake, Qld, a bird gave 117 Territorial Songs in 55 min of recorded Song, including six segments of Gronking Song, but no Clicking. In late June, at Lamington NP, Qld, in June, a bird gave 38 Territorial Songs, then, in 55 min, 60 cycles of Sequential Song interspersed with 44 Territorial Songs, first thing in the morning.
- Subsong: After the breeding season, with the onset of moult, start to give quiet Subsong,which includes much mimicry, as well as a subdued version of Lyrebird vocalizations; some mimicry of man-made sounds may occur (Robinson 1975; see above). Subsong does not stop with the end of moult, but gradually merges into full Song with the start of the male gonadal cycle (Robinson 1975). Subsong reported in mid-Sept. (V. Powys).
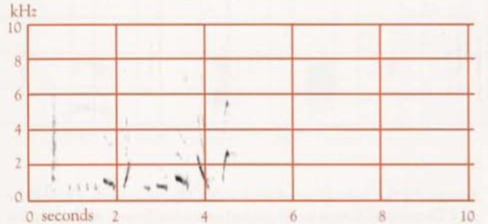
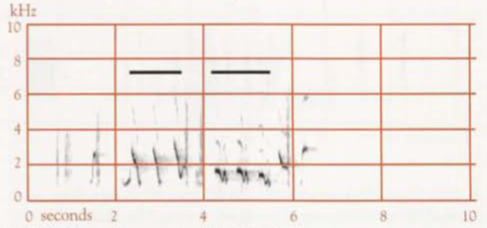
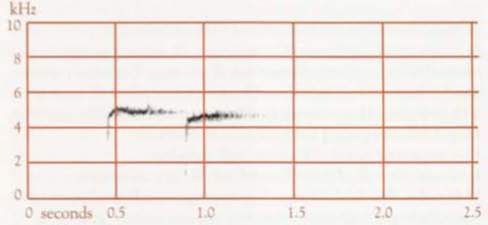
- Territorial song: One to several very short introductory notes, followed by a short pause, and then a series of mostly longer notes; duration of the whole mostly 5–10 s (Robinson & Curtis 1996). An earlier study found that, including the usual 1–3 introductory notes, duration varies from 3.5 s to 5.6 s, and frequency from 0.5 to 3.6 kHz (Reville 1973). The main body of the Song contained from 5 to 11 notes, of which only five or six might be audible >30 m from the bird (Reville 1973), and of which the last is usually a high frequency, ascending element (Backhouse et al 2021). The harmonic structure gives a clear, almost brilliant, quality (Reville 1973). The bird typically pauses briefly after each Territorial Song. If the Song of another male is audible during this pause, the bird often waits until the other is finished, then either repeats the introductory notes and gives the full Territorial Song or abandons the attempt and continues with Sequential Song. For discussion of mimicry in the Territorial Song, see above. Mosaic geographical variation occurs, with one, two, and sometimes three Territorial Song-types present in a population, which in basic pattern are shared by all members of that population (Reville 1973). Variants in the Song-types involve little more than the insertion, deletion or transfer in position of minor figures (Reville 1973), e.g. omission or changing of introductory notes, addition of notes in the middle, or, more rarely, at the end (Robinson & Curtis 1996). An individual may do this on one occasion, yet on others produce the typical form of the song type (Reville 1973). There is one type of Territorial Song at the Knoll Section of the Tamborine Mountain National Park, Qld (sonagram A); three at Lamington National Park, Qld (one shown in sonagram B); and six at Toonumbar State Forest, NSW, (two shown in sonagrams C and D; in the latter the horizontal bars indicate mimicry of the Satin Bowerbird, left, and Pied Currawong, right).
- Sequential song (=Albertcycle Song, Display Song, Recital Mimicry): A stereotyped sequence, consisting of mimicry of songs and calls of local bird species and sounds of bill-clicks, wings, large bird landing on branch, etc., lasting 40–50 s, and repeated over and over. At any one location, 3–10 species of bird, and up to 37 different sounds, are mimicked (Backhouse et al 2023). Sonagram E shows four sequences.Mostly much softer than Territorial Song. All males in a particular locality use a similar pattern of sounds, with sequences almost as similar between individuals as within individuals (Backhouse et al 2022). In the only area where such a study was made, the pattern appeared unchanged after 19 years. Repetition is not perfect: sometimes one or a few components are missed, or a component is varied, e.g. one note given instead of the usual three. Incomplete sequences often occur. Often a bird interrupts Sequential Song to listen to Territorial Song of a distant male, and sometimes responds with Territorial Song. He then continues from where he had stopped in the Sequential Song, starts again from the beginning, or simply omits the sounds that he would have given in the time he was uttering Territorial Song or listening. Less often, he starts in a different part of the Sequential Song altogether.
- Gronking song: Repeated short loud gronk notes, which carry well (‘Loud Gronking’: Backhouse et al 2024), preceded and followed by softer sounds audible only at close range. The preceding sounds are softer crackling and loud cracking sounds; the sounds that follow are a series of very soft non-musical notes (‘Regular Gronking’: Backhouse et al 2024), usually at two different low pitches, forming a regular rhythmic pattern of 2–6 ‘beats to a bar’ (the number of beats being constant at any one locality), but sometimes the series consists of a single first note and a second note repeated 10–13 times (H.S. Curtis). Usually the same pattern of rhythmic notes is used by all birds at a particular locality. Sonagram F shows three gronks, then rhythmic notes at three ‘beats to the bar’ (these notes are softer and have been amplified), and finally three more gronks. Within a Song performance, sometimes only gronks alone are given, but more usually the Gronking Song is given once or several times, usually ending with gronks, before returning to the Sequential and Territorial Songs. Harsh sounds, in some cases mimicry of a brushtail Trichosurus possum, immediately follow the Gronking Song, in turn often succeeded by mimicry of the loud musical song of Grey Shrike-thrush or song of Eastern Whipbird. Gronking Song does not occur in all sessions of Song but does always occur when a female visits the display platform (F. Backhouse). It may also be heard independently of Song, probably away from the display platform. Gronking Song is possibly analogous to the plik songs (or ‘Dance Song’: Dalsiell et al 2013) of Superb Lyrebird (Curtis 1998; V. Powys; F. Backhouse).
- Clicking: Soft clicks, easily overlooked, and heard only rarely. Recorded immediately after copulation (F. Backhouse). One bout of Clicking lasted 20 s. Probably analogous to Clicking of Superb Lyrebird (Curtis 1998).
- Intense alarm call: A piercing shriek (two shown in sonagram G), similar to that of the Superb Lyrebird (Keast 1944; Robinson & Curtis 1996); rendered whisk-whisk (Keast 1944). The loud sudden whistle of Jackson (1921) is probably this call.
- Other alarm calls: Other calls apparently used in threat and alarm are many, varied, and often harsh; at Toonumbar Sate Forest, include mimicked calls of Eastern Whipbird (but not its loud antiphonal song).
- Other calls: Male uttered slow clucking, that become faster during copulation; afterwards, he continued clucking softly (Smith 1988). In Nov., in an encounter between a male and a smaller bird, apparently also a male, the larger uttered loud single quilp calls followed by double calls of quilp, quilp (Clarke 1968).

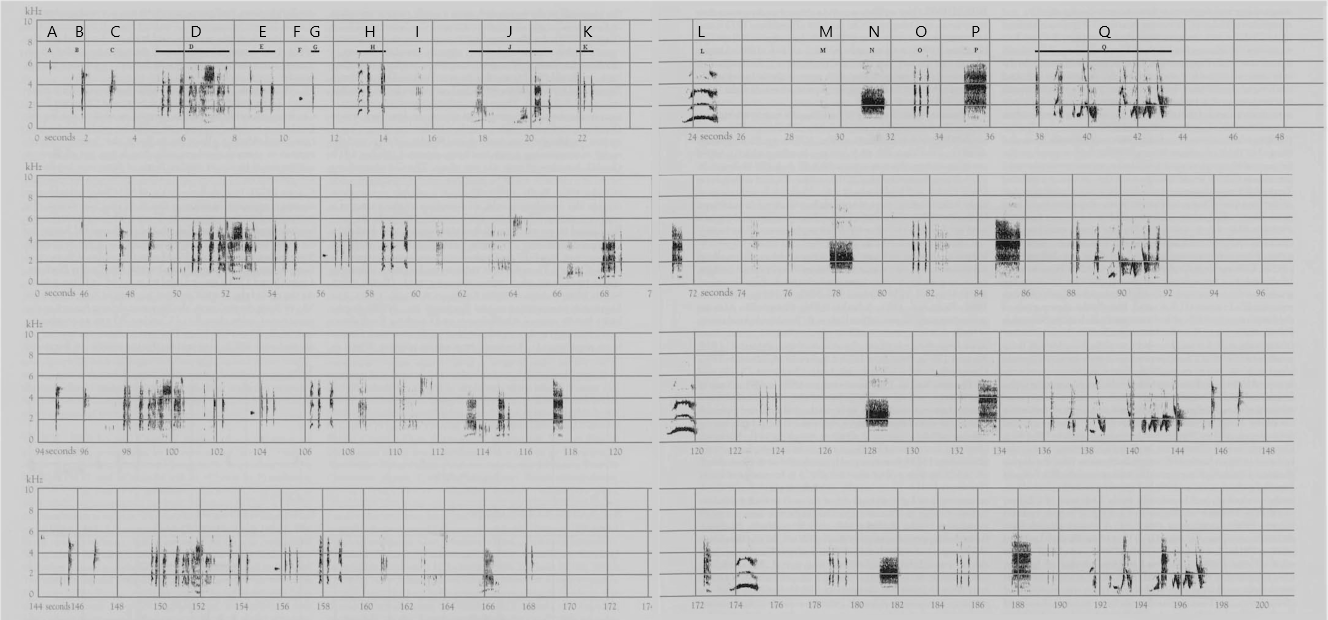
A Very short high-pitched note of Green Catbird;
B Australian King-Parrot;
C Australian Logrunner;
D series of mostly non-musical sounds of Satin Bowerbird;
E bill taps;
F musical piping note of Eastern Yellow Robin;
G non-musical notes of Eastern Yellow Robin;
H non-musical alarm notes of Lewin’s Honeyeater;
I rapid bill snapping;
J Laughing Kookaburra;
K non-musical notes of Eastern Yellow Robin;
L advertising call of Satin Bowerbird;
M flapping wings;
N Paradise Riflebird;
0 sounds not recognised;
P harsh note of Satin Bowerbird;
Q series of mostly musical sounds of Satin Bowerbird.

Adult female
Rarely call close to nest, except when flushed or when young threatened (L. Nielsen).
- Song and mimicry: Capable of both (Curtis 1998). One female at Tamborine Mountain was recorded singing in response to the recordist disturbing her while carrying nesting material, with mimicry of Grey Goshawk Accipiter novaehollandiae, Crimson Rosella Platycercus elegans, Satin Bowerbird Ptilonorhyncus violaceus and Eastern Whipbird Psophodes olivaceus, and Song reminiscent of the local male Territorial Song (Backhouse 2021). Another female at Goomburra in Main Range National Park with a nestling was recorded near the nest mimicking the calls of Eastern Whipbird and Grey Goshawk, along with own alarm calls. The mobbing calls of several other species in the recording suggest the presence of a predator near the nest (Backhouse 2021).
- Intense alarm call: Loud piercing single-note scream uttered when flushed from nest while incubating (Beruldsen 1973; L. Nielsen).
Young
Little information. Chick responded with screech to an alarm call from its mother (H.S. Curtis). Claim that males do not utter Song until fourth year (Gould) considered doubtful (Smith 1988). Thought that mimicked calls learnt from adult males (see above).
Breeding ⓘ
Not well known and no major studies; some data from nests at Dalrymple Creek, south-eastern Qld (Beruldsen 1973); and observations at one nest at O’Reilly’s, Lamington National Park, south-eastern Qld (Lindsey 1994); four records in NRS to Feb. 1998. Mating system not known (see Social Organization). At Tamborine Mountain, some birds can go for up to 5 years without breeding (L. Nielsen). Almost no breeding in years of severe drought (H.S. Curtis; L. Nielsen).
Season
Eggs, late May to late-Aug. (Keast 1944; Nielsen 1959a; Beruldsen 1973; Campbell; North; NRS; G. Holmes; L. Nielsen; F. Backhouse & A. Maisey). One nest in Blackwall Range., NSW: egg, 30 May; small nestling, a few days old, 15 July; large nestling, which jumped out of nest when observer approached, 12 Aug. (G. Holmes). In 1973, laying began late May on Tamborine Mountain, and early July on Great Divide and McPherson Ranges. (L. Nielsen). Female on nest, early June; nestling, Sept. (Aust. Atlas); fledging to mid-Sept. (Schodde 1996). Campbell claims a heavily incubated egg in a nest in Oct. but no other reference supports such late laying.
Site
Nest beneath canopy of forest (see Habitat). On rock-ledges, in clefts in rocks, or between two rocks, occasionally in caves; on rocky cliff-faces, rock-faces on steeply sloping ridges, or in deep rocky ravines; sometimes near waterfalls (Favaloro 1931; Marshall 1935; Keast 1944; Nielsen 1959a; Chisholm 1967; Beruldsen 1973; Schodde 1996; Campbell). Also on ground on steep slopes: on bank of creek, between buttress roots of fig Ficus, among stumps, at base of palm, on rock, among ferns or in dense shrub; occasionally in fork of tree; (Marshall 1935; Nielsen 1959a; Beruldsen 1973; Lindsey 1994; Schodde 1996; Campbell; North; NRS; L. Nielsen). On Great Divide and McPherson Ranges, south-eastern Qld, most sites (c. 65%; no N) at base of small tree, often a Bangalow Palm Archontophoenix cunninghamiana, and often in a patch of Rainforest Spinach Elatostema reticulatum, within rainforest on steep hillsides; on Tamborine Mountain, of ten nests, all were on rock-faces (L. Nielsen). One nest in centre of bird’s-nest fern, in upright fork of tree (Beruldsen 1973); one nest partly concealed with plant debris (North); one nest at foot of waterfall c. 6 m from edge of water (Favaloro 1931). Nests usually in darkest part of rainforest; occasionally in light, in high situation such as rock-face close to top of ridge (L. Nielsen). In Dalrymple Creek area, all nests found were on the shaded n. side of E–W valley (L. Nielsen). Recorded aspects of entrance: SE × 2, SW × 1, N × 1 (Beruldsen 1973), S × 1, W × 1 (F. Backhouse & A. Maisey); aspect of nest related to features of site, female requiring easy access and ability to glide away from nest (L. Nielsen). Two active nests c. 80 m apart (L. Nielsen). May re-use same site, but only after old nest has disintegrated such that only platform remains (L. Nielsen). A female can sometimes be found nesting within 60 m of nest of a previous year; one female (identified as the same female by her unusually marked egg) nested within an area of 50 m diameter over a 10-year period (L. Nielsen).
Measurements: Height of nest: in cliff or rock-face, 4.9 m (2.64; 1–9; 7) (Nielsen 1959a; Beruldsen 1973; NRS); in fork of tree, 2–5 m (no N) (Schodde 1996). Dimensions of one rock-ledge: width, 100 cm; depth, 60 cm (Beruldsen 1973).
Nest, Materials
Build dome-shaped nest with side entrance; composed of sticks, fern fronds, rootlets, pieces of palm leaf, and moss; lined with moss, fine plant material and feathers (Nielsen 1959a; Beruldsen 1973; Campbell; North; NRS). Of four nests at Dalrymple Creek, three were a solid platform of short dry sticks, built up on each side to form partial walls; the fourth was just a rudimentary flat platform of sticks; on each platform was built a neat rounded dome, constructed from blackish fern rootlets, with some plant fibre and bark; roof of some domes may protrude over entrance, forming a small hood; inside of nest-cavity, smooth at sides, with no loose ends protruding, and often lined with feathers (Beruldsen 1973). Nests in Great Divide and McPherson Ranges extremely neat, with well-constructed domes; nests at Tamborine Mountain crude, with roughly constructed domes, with one so poorly constructed that part of the nest had fallen away, taking the egg with it (L. Nielsen). Cavity of two nests, early in incubation period, contained 27 and 68 Lyrebird feathers; nests may contain over 150 feathers towards end of incubation period (Beruldsen 1973). Egg hidden from view by feathers lining nest (Beruldsen 1973), though North said egg in one nest visible through entrance. Only female builds (Schodde 1996). Construction of nest may take at least 3 weeks (NRS). Young non-breeding females may construct a nest-platform with partial walls (Beruldsen 1973).
Measurements: One nest (cm): length × width × depth of platform, 67.5 × 52.5 × 2.5; length × width × height of exterior of dome, 35 × 35 × 35; length × width × height of interior of dome, 25 × 30 × 26.3; horizontal × vertical diameter of entrance, 15 × 11.3 (Beruldsen 1973). Two nests in Goomburra (Main Range National Park) had the following measurements: one nest (cm): length × width × height of exterior of dome, 60 × 52 × 45; length × width × height of interior of dome, 37 × 29 x 30; horizontal × vertical diameter of entrance, 19 × 11; other nest (cm): length × width × height of exterior of dome, 30 × 57 × 45; length × width × height of interior of dome, 34 × 30 x 32; horizontal × vertical diameter of entrance, 17 × 14 (F. Backhouse & A. Maisey).
Eggs
Oval to short oval; ground-colour, pale vinous-brown, dull dark olive-brown or purplish brown, uniformly marked with spots, blotches and, sometimes, short streaks, of brown to blackish brown, larger and more numerous on larger end, forming an irregular band, and in some places, confluent patches; some markings dull and scarcely distinguishable from ground-colour (North). Longish oval; coarsely textured, pitted and uneven; slightly glossy; ground-colour, purplish grey to dark purplish grey, sparingly blotched with dark brown or sepia, and dull or cloudy markings, mostly at larger end (Campbell). Lustreless when freshly laid, becoming polished and partly glossy during incubation; ground-colour, dull darkish grey, with or without slight brownish or purplish tinge; one sparsely and more-or-less evenly spotted with dull blackish grey; another sparsely spotted but with cap of blackish grey over larger end (Beruldsen 1973). Eggs from Great Divide and McPherson Ranges vary greatly in colour and, sometimes, shape; eggs from Tamborine Mountain nearly identical in colour and shape (L. Nielsen).
Measurements: 61.8 (2.21; 59.4–64.5; 6) × 43.1 (0.70; 42.2–43.7) (Campbell; North).
Clutch-size
One (Storr 19; Campbell). Laying Egg may be laid up to 1 month after completion of nest (NRS). Said to raise only one brood per season (Schodde 1996) but basis for statement not known.
Incubation
By female only (Keast 1944; Nielsen 1959a; Beruldsen 1973; Lindsey 1994; Schodde 1996). Female usually leaves nest at c. 09:00 and returns at c. 15:00; during period of absence egg becomes quite cold (L. Nielsen).
Young, Growth
No information.
Parental care, Role of sexes
Only female feeds and broods young (Lindsey 1994; Schodde 1996). When brooding, one female sat facing entrance of nest, with tail curved up over her back and protruding out of entrance (Lindsey 1994). Female removed faecal sacs (Lindsey 1994).
Fledging to independence
Fledging period: c. 5.5 weeks (Lindsey 1994). No other information.
Success
No information.
Plumages ⓘ a-z
Prepared by A.M. Dunn. Fledge in juvenile plumage. Early plumages not well understood and sequence to adult plumage not known. After attaining adult plumage, complete post-breeding (pre-basic) moults each cycle produce successive adult plumages without change in appearance. In adult and immature plumages, sexes differ only in structure, pattern and colour of tail; sexes alike in juvenile plumages. No subspecies. Descriptions based on examination of skins of 17 adult males, nine adult females, two immature males, and two juveniles.
Adult male
(Definitive basic).
- head and neck: Forehead, dark brown (c121) with red-brown (c32) to dark red-brown (132) suffusion. Crown, nape and hindneck, dark brown (c121). Ear-coverts, dark brown (121) with a red-brown (c32) suffusion. Chin, throat, foreneck and sides of neck, rufous-brown (c240) with light grey-brown (119D) bases to feathers. Lores and area surrounding eye, very sparsely feathered with small dark-brown (c121) feathers.
- upperparts: Mantle, back, scapulars and uppertail-coverts, dark red-brown (ne) with concealed, downy, light grey-brown (119C) bases. Rump slightly paler than rest, rufous-brown (c36).
- underparts: Most of breast and belly, light grey-brown (c119D); upper breast has buff (124) to buff-brown (c39) suffusion to tips of feathers; and grey-brown grades into light rufous (ne) on sides of lower breast. Flanks grade from light grey-brown (c119D) anteriorly, through light rufous (ne), to rufous-brown (c36) posteriorly. Lower belly, pale grey (c86). Feathers of vent, down-like, with rufous-brown (c36) tips and brown (c28) bases. Undertail-coverts, red-brown (132A).
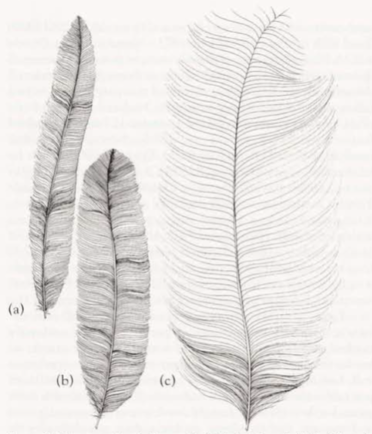
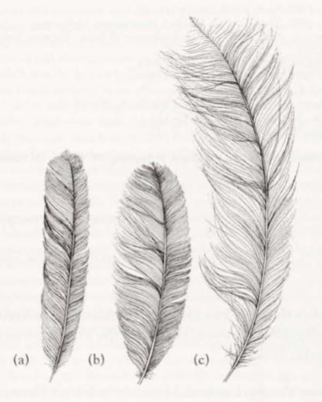
- uppertail: Rectrices modified and used for highly elaborate display. Rectrices can be divided into three types (following terminology of Smith 1982): medians (t1), filamentaries (t2–t7) and lyrates (t8) (see also Superb Lyrebird). Medians (t1), long and very slender (c. 15 mm wide) with strong curve toward inner web near tips; outer webs have very reduced barbules; medians, dark brown (219A), usually with a rufous-brown (c36) suffusion near rachis. Filamentaries (t2–t7), long and very broad (c. 150 mm wide) with much reduced barbules on both inner and outer webs of distal half of feathers; all filamentaries have a slight concave curve to the dorsal surface, causing feathers to curve upward. Lyrates (t8), long and broad (c. 100 mm wide), but shorter than other rectrices, and show little reduction in barbules. Filamentaries and lyrates (t2–t8), black-brown (119).
- undertail: Medians (t1), brownish grey (c79). Filamentaries and lyrates (t2–t8), dark grey (83) with white rami and light grey-brown (119D) shafts; white rami may catch light under some light conditions, resulting in paler silvery appearance.
- upperwing: Marginal and median secondary coverts, dark red-brown (ne). Greater secondary coverts, tertials and secondaries, dark red-brown (ne) on outer webs and black-brown (c119) on concealed inner webs. Marginal primary coverts, brownish grey (c79). Lesser and median primary coverts, dark brown (c121). Greater primary coverts, dark brown (c21). Feathers of alula, black-brown (c119) at base, grading to dark brown (c21) in middle of feathers, to light brown (25) at tips. Primaries, dark brown (c121) on outer webs and black-brown (c119) on concealed inner webs.
- underwing: Marginal, lesser and median coverts, brownish grey (c79). Greater coverts, grey (c84). Tertials, secondaries and primaries, dark brownish grey (brownish 83).
Adult female
(Definitive basic). Similar to adult male, differing only in tail, and then mainly in structure (see Structure for differences) and only slightly in coloration:
- uppertail: Medians (t1), long and relatively slender (c. 40–50 mm wide) with tapered tip, but still much broader and shorter than in adult male; medians, dark brown (121) with strong rufous-brown (c36) suffusion, which is strongest on distal half of feather and near rachis. Filamentaries (t2–t7) and lyrates (t8), long and broad (c. 60–70 mm wide) with rounded tips and no reduction in barbules; shorter and narrower than those in adult male. Coloration of filamentaries as adult male, but with red-brown (c32) suffusion near tips; extent of suffusion reduced toward outer rectrices, and confined to diffuse fringe on t7. Coloration of lyrates as adult male but with narrow diffuse red-brown (c32) fringe to tips.
- undertail: Dark grey (83) to grey (84) with weak light brown (c223C) suffusion to tips of inner rectrices.
Nestlings
No observations of hatchlings. Older nestling, c. 40 days old (QM), covered with thick down on upperparts and underparts, but developing juvenile plumage on head, neck and wings. Down of upperparts, dark brown (grading from 121A on mantle to 121 on rump). Down of underparts, dark brown (219A) on upper breast, light grey-brown (c27) on lower breast and flanks, off-white (ne) to pale grey (86) on belly and vent, and grey (84) on thighs. Head and neck mostly bare, with juvenile plumage on forehead, crown and nape. Wings covered in developing juvenile plumage. Tail not yet developed.
Juvenile
Plumage of head, neck, body and wings similar to adult but with slight differences; tail differs in structure from adult male and female, but similar in coloration to tail of adult female.
- head and neck: Forehead and forecrown, redder than in adult, red-brown (c32). Rest of crown, and nape and hindneck slightly lighter than in adult, brown (28). Ear-coverts, red-brown (c221B). Chin, throat, foreneck and sides of neck, rufous-brown (c240), appearing more uniformly rufous than in adult. Rest as adult.
- upperparts: Mantle, back, scapulars and uppertail-coverts similar to those of adult but slightly paler. Feathers of rump soft and down-like, light rufous-brown (c139).
- underparts: Breast, orange-rufous (c132D). Feathers of flanks, down-like and light rufous-brown (c139). Feathers of most of belly, vent and undertail-coverts, down-like and pale grey (c86); those of upper belly and undertail-coverts have light rufous-brown (c139) wash.
- tail: Medians (t1) similar in shape to those of adult female. Filamentaries (t2–t7) and lyrate (t8) similar to those of adult females in length, but much narrower (c. 35–50 mm wide) and more tapered toward tips; no reduction in barbules. Coloration similar to that of adult female, but with brown (c121B) suffusion to basal half of rectrices and weak light rufous-brown (c139) to rufous-brown (c136) suffusion to tips of all rectrices; suffusion diffuse and not confined to fringe on outer rectrices (cf. adult female).
- wing: As adult male.
Immature male
(First and unknown number of subsequent basic plumages). Little information available. Younger males appear as adult in plumage of head, neck, body and wings, but differ in structure and coloration of tail. Medians (t1) similar to those of adult female in younger males; probably become gradually narrower and longer after each moult till adult male plumage eventually attained. Filamentaries (t2–t7) of younger males mostly black-brown (119), merging into broad rufous-brown (136) tips, which probably lost gradually with increasing age; in younger males, most filamentaries similar in structure to those of adult female, but some have reduced barbules and widely spaced rami near tips so that tips have similar appearance to the filamentaries of adult male; the number and proportion of modified filamentaries probably increases with age till adult male plumage eventually attained. Lyrates (t8), black-brown (119) with rufous-brown (136) suffusion at tip, that probably lost gradually with age. Age of maturity not known, but probably similar to that of Superb Lyrebird, which is c. 8 years.
Immature female
(First and unknown number of subsequent basic plumages). None identified among skins. Probably as adult female.
Bare Parts ⓘ a-z
Based on photos (Lindsey 1994; and standard sources) and museum labels (QM).
Adult
Bill, black (89). Iris, black (89); or dark brown (QM). Inside mouth, dark grey (QM). Orbital ring, dark blue-grey (78) or light blue-grey (c88). Legs and feet, brownish grey (79), dark grey (83) or black (89). Claws, dark grey (83).
Nestlings
From photos of older, mostly feathered nestling (Lindsey 1994): Bill, mostly grey-black (82) with pale grey (86) tip, and grading to light grey (85) at base of lower mandible. Gape, grey (c84). Bare skin on face and orbital ring, grey (c84). Legs, dark brown (c121). Feet, grey-black (c82).
Juvenile
No information.
Immatures
As adult.
Moults ⓘ
Based on examination of 15 adult skins (ANWC, HLW, QM, SAM). Very poorly known.
Adult post-breeding
(Pre-basic). Few data; available skins were collected Feb. (n=1) and May–Sept. Probably complete, and primaries outward. None had active moult of primaries. Only two had active moult of tail: one in Feb. was moulting t1 and t3 on right side, and one in May had heavy moult of tail on one side. One in Feb. had slight moult of body, on head, neck and underparts, and one in July had slight moult of upperparts. No others were recorded with active moult. Adult males in Lamington National Park observed on trail cameras with partial tail moult from late September to January, though none observed with all tail feathers missing as is common in superb lyrebirds (F. Backhouse).
Post-juvenile
(First pre-basic). No information.
First and subsequent immature post-breeding
(Second and subsequent pre-basic). No information.
Measurements ⓘ a-z
Measurements from skins of adult and juvenile birds from ne. NSW and se. Qld (AM, ANWC, HLW, MV, QM, SAM):
- Wing
| Age | Males | Females | Significance |
|---|---|---|---|
| Adult | 275.5 (6.97; 265–287; 17) | 259.2 (13.31; 240–276; 9) | ** |
| Juvenile | – | 252 |
- Tail median (length of longest median [t1])
| Age | Males | Females | Significance |
|---|---|---|---|
| Adult | 540.1 (27.34; 490–585; 14) | 392.2 (19.34; 360–423; 8) | ** |
| Juvenile | – | 428 |
- Tail lyrate (length of longest lyrate [t8])
| Age | Males | Females | Significance |
|---|---|---|---|
| Adult | 379.1 (21.75; 348–425; 15) | 264.0 (16.38; 246–286; 6) | ** |
| Juvenile | – | 256 |
- Tail filamentary (length of longest filamentary [t2–t7])
| Age | Males | Females | Significance |
|---|---|---|---|
| Adult | 530.1 (29.11; 469–572; 15) | 342, 356, 370 |
- Bill S (chord of the culmen from the bill tip to the naso-frontal hinge)
| Age | Males | Females | Significance |
|---|---|---|---|
| Adult | 40.8 (2.56; 34.3–44.3; 16) | 40.7 (3.00; 37.8–45.6; 7) | ns |
| Juvenile | – | 37.4 |
- Tarsus
| Age | Males | Females | Significance |
|---|---|---|---|
| Adult | 110.7 (3.91; 104.9–116.6; 16) | 102.9 (6.64; 96.1–116,7; 9) | ** |
| Juvenile | – | 96.1 |
- Length of middle toe (Toe C)
| Age | Males | Females | Significance |
|---|---|---|---|
| Adult | 81.2 (3.61; 76.8–86.9; 9) | 78.6 (4.70; 73.0–84.6; 6) | ns |
| Juvenile | – | 74.6 |
Weights ⓘ
Weights of NE. NSW and se. Qld adults, from museum labels (ANWC, QM):
| Males | Females |
|---|---|
| 927.8 (240.84; 668–1215; 6) | – |
Nesting female caught at Goomburra (Main Range National Park) weighed 973 g (F. Backhouse & A. Maisey).
Structure ⓘ a-z
Wing short, broad and rounded. Eleven primaries (Morlion 1985; this study): p1–p3 longest (p4–p6 sometimes equal longest); p10 92–120 mm shorter, p9 47–63, p8 24–37, p7 5–17, p6 0–5, p5 0–3, p4 0–3; p11 minute. Primaries not emarginated. Fourteen secondaries (Morlion 1985; this study), including about three tertials; tips of longest tertials fall short of secondaries on folded wing.
Tail: Differs between sexes and between ages, with considerable structural modification to rectrices of adult male and with little or no modification to rectrices of females and young males; 16 rectrices. In adult male, tail long and broad and composed of three different types of modified feathers: medians, filamentaries and lyrates. In adult female, tail long, but much shorter than male, with rather plain rectrices. See Plumages for detailed descriptions of each of the rectrices (also see Figs 2–4).
Bill narrow and pointed, about same length as head; upper mandible curves slightly to pointed tip; lower mandible rather straight and flat with shallow taper to pointed tip. Nostril long and oval shaped (c. 7 mm), and set in large shallow nasal groove.
Tarsus long and thick; scaling, scutellate. Tibia fully feathered. Outer toe 91–97% of middle, inner 93–97%, hindtoe 80–89%.
Ageing, Sexing ⓘ
No information in HANZAB books.
Recognition ⓘ
No information in HANZAB books.
Geographical Variation ⓘ
None, although differences in aspects of plumage, egg colouring and structure of nest have been noted from the Tamborine Mountain metapopulation, isolated for some time from the Lamington Plateau (L. Nielsen, pers. comms.)
References (see also Simplified Ref.)
- Backhouse, F. 2021. PhD thesis, Western Syd. Univ., Sydney.
- ——, et al. 2021. Ecol. Evol. 11: 2701-2716.
- ——, et al. 2022. Proc. R. Soc. B. 289(1970): p.20212498.
- ——, et al. 2023. Divers. Distrib. 29(1): 109-122.
- ——, et al 2024. AmNat, in press.
- Behrens, D. 1994. The Elusive Rainforest Troubadour. [Video]. Author, Rochedale, Qld.
- Beruldsen, G.R. 1973. Aust. Bird Watcher 5: 44–8.
- Brouwer, J., & S. Garnett. 1990. RAOU Rep. 68.
- Chisholm, A.H. 1946. Nature’s Linguists. Brown Prior Anderson, Melbourne.
- —— 1957. Emu 57: 25–30.
- —— 1960. Emu 60: 145–6.
- —— 1967. Emu 66: 373–6.
- Clarke, J.H. 1968. Qld Nat. 15: 105–6.
- ——, & C. Clarke. 1972. Sunbird 3: 76–9.
- Curtis, H.S. 1972. Emu 72: 81–4.
- —— 1998. Nature Aust. 26(1): 32–41.
- Favaloro, N.J. 1931. Emu 31: 48–59.
- Garnett, S. (Ed.) 1993. RAOU Rep. 82.
- Gosper, D.G. 1986. Corella 10: 1–16.
- Hartshorne, C. 1953. Emu 53: 109–28.
- —— 1973. Born to Sing. Indiana Univ. Press, Bloomington, USA.
- Holmes, G. 1987. Avifauna of the Big Scrub Country. NSW Nat. Pks. & Wildl. Serv., Sydney.
- Irby, F.M. 1930. Emu 29: 276–7.
- Jackson, S.W. 1909. Emu 9: 45.
- —— 1920. Emu 19: 258–72.
- —— 1921. Emu 20: 195–209.
- Keast, J.A. 1944. Emu 43: 177–87.
- Lawler, W. 1980. Wildlife (Lond.) 22(9): 44–6.
- —— 1981. Wildl. Aust. 18(2): 50–3.
- Lindsey, T. 1994. GEO A’asia 16(4): 104–11.
- Macdonald, J.D. 1969. Emu 69: 118–19.
- Mack, G. 1952. Victorian Nat. 69: 68–70.
- Marshall, A.J. 1935. Emu 35: 36–48.
- Mitchell, K. J., et al. 2021. Biol. J. Linn. Soc.134(1): 40-56.
- Morlion, M.L. 1985. Rec. Aust. Mus. 37: 143–56.
- Morris, A.K., et al. 1981. Handlist of Birds in New South Wales. NSW Field Orn. Club, Sydney
- Nielsen, L. 1959a. Aust. Bird. Watcher 1: 51–2.
- —— 1959b. Emu 59: 249.
- —— 1960. Aust. Bird. Watcher 1: 109–12.
- Putland, D.A., et al. 2006. Biology Letters 2(3): 367-370.
- Ratcliffe, F. 1938. Flying Fox and Drifting Sand. Chatto & Windus, Lond.
- Reilly, P.N. 1988. The Lyrebird. Univ. NSW Press, Sydney.
- Reville, B. 1973. Unpubl. BSc (Hons) thesis, Univ. Qld, Brisbane.
- Roberts, G.J. 1977. Sunbird 8: 73–82.
- —— 1979. The Birds of South-East Queensland. Qld Cons. Council, Brisbane.
- Roberts, S. 1930. Emu 29: 195–200.
- Robertson, J.S. 1948. Emu 47: 373–88.
- ——, & A.R. McGill. 1948. Emu 47: 373–88.
- Robinson, F.N. 1974. Emu 74: 9–10.
- —— 1975. Emu 75: 23–7.
- —— 1991. Emu 91: 61–3.
- ——, & H.S. Curtis. 1996. Emu 96: 258–75.
- Schodde, R. 1996. Murwillumbah Management Area Fauna Survey 1996. CSIRO Div. Wildl. Ecol., Canberra.
- Smith, A.P., et al. 1994. Terrestrial Fauna of the Grafton and Casino State Forest Management Areas. Description and Assessment of Forestry Impacts. Grafton–Casino Management Area Environmental Impact Statement Supporting Document 1. Report prepared by Austeco Consultants for State Forests of NSW, Sydney.
- Smith, L.H. 1968. The Lyrebird. Lansdowne Press, Melbourne.
- —— 1982. Aust. Wildl. Res. 9: 311–30.
- —— 1988. The Life of the Lyrebird. Heinemann, Melbourne.
- Stewart DA, Legge SM, Ehmke G, Garnett ST (2021) Albert’s Lyrebird Menura alberti. In The Action Plan for Australian Birds 2020. (Eds ST Garnett and GB Baker) pp. 472–474. CSIRO Publishing, Melbourne.
- Thomson, A.L. 1964. A New Dictionary of Birds. Thomas Nelson, Lond.
- Trudgeon, M. 1969. Aust. Birds 4: 9–11.
- —— 1973. Birds (J. NSW FOC)8: 45–6.
- Vidler, E.A. 1933. Emu 33: 132.
- Wheeler, W.R. 1973. Aust. Bird Watcher 4: 257–69.
- White, S.A. 1910. Emu 10: 212–15.
- Wigan, M.L., et al. 1938. Emu 37: 25–7.
Bibliography
Original Text from HANZAB book
View PDF in browser or Download PDF to open outside your browser.
Note that these links may not work as described, with the behaviour dependent on the browser being used and your settings for viewing PDFs.
Original HANZAB Sponsors: HS Curtis, BR Grant, P Heyning, FF Whyte, JN Yates
Updated 18 February 2025
Multimedia
Links to external multimedia. The linked sites may not use the same taxonomy as HANZAB. Some multimedia items may also be included in the entry, such as audio in the Voice section.
Photographs
Videos
Audio
. . . . .
Content Updates
(For site wide changes and release dates, see Release Notes)
Conservation Status addedMap updated
Release 1.2
- description of the last revision (see below) updated with the sections that have changed
- accessibility changes to headings and tables
Release 1.1
- Text revised by F. Cehak & F. Backhouse. Sections changed (other than minor text changes):
- Field Identification (introduction; Similar species)
- Distribution and Population (Change in range, populations)
- Threats and Human Interactions
- Movements (Banding)
- Social Organisation (introduction; Bonds; Size of territories)
- Social Behaviour (Display with tail; Copulation)
- Voice
- Breeding (Season; Site; Nest Materials,Measurements)
- Moults (Adult post-breeding )
- Weights
- References
- abbreviations have been removed eg ne change to north east
Recommended Citation
F. Cehak & F. Backhouse (2025) Albert’s Lyrebird (Menura alberti). In Handbook of Australian, New Zealand and Antarctic Birds, published on 1.1. HANZAB Release ? 2026. Birdlife Australia. [Text before updates sourced from: Marchant, S. et al (eds) 1990-2006 Handbook of Australian, New Zealand and Antarctic Birds.Volume 1 to 7. Birdlife Australia.] Source: https://hanzab.birdlife.org.au/species/alberts-lyrebird/ Release ? 2026. Accessed: April 29, 2026 Time Zone: +10:00
. . . . .