This document is designed to assist readers in understanding the information presented in HANZAB. It consolidates the explanatory information from the introductions of all seven volumes (for the original text, see introductions for Vol. 1, 2, 3, 4, 5, 6, 7) along with information about changes in presentation in the HANZAB website. For lists of the species included in each volume of the HANZAB books see the contents for each volume: 1, 2, 3, 4, 5, 6, 7.
NOTE: Threats and Human Interactions was not included as a separate section in volumes 1 to 4 of the books. For consistency, this heading has been added to these entries and links to relevant text within the entries created. Conservation Status has also been added. New distribution maps have been added to some species but links to the original maps have been included below the new map. Research published after the HANZAB book are included in the Bibliography.
Website presentation changes
The move to a website has allowed a number of changes to the presentation:
- some paragraphs have been split to make them more readable.
- reference below to first paragraph, second paragraph etc may not be correct
- hyperlinks have been added
- links to external site annotated with
- hyperlinks to broken links being removed but text retained (process of identifying broken links underway – R2.0)
- Other English names altered to better support search by fully stating species names rather than grouping them
- taxonomic changes and changes to the content of entries documented
TAXONOMY AND NOMENCLATURE
The taxonomy has been updated using AviList 2025 for orders, families and species. Subfamilies are not included in AviList 2025 but are flagged for future releases. Subfamilies have been retained as per the HANZAB 2023 but see notes in the family entry for issues raised by changes to the taxonomic sequence of species (eg Anatidae). Subspecies, although included in AviList 2025, have not been finalised. Species split or merged in AviList 2025 have the subspecies changed. Notes about inconsistencies at the subspecies level are included in the species Taxonomic Changes.
Nomenclature of orders, families and species in the HANZAB books has been updated using the following sources:
- AviList 2025
- Working List of Australian Birds (Birdlife Australia Version 4.1 and 4.3), referenced as WLAB 4.1 and WLAB 4.3;
- Checklist of the Birds of New Zealand, Fifth Edition, 2022 (Ornithological Society of New Zealand) referenced as NZCL 2022;
- Birdlife International datazone website for species recorded in Antarctica, French Southern territories, Heard and MacDonald islands, Bouvet Island, and South Georgia and South Sandwich Islands (Accessed 10 June 2025); and
For HANZAB 2023 the following sources were also used:
- Birdlife International datazone website for species recorded in Antarctica, French Southern territories, Heard and MacDonald islands, Bouvet Island, and South Georgia and South Sandwich Islands (Accessed 23 March 2023); and
- for extralimital subspecies, HBW and BirdLife International (2022) Handbook of the Birds of the World and BirdLife International digital checklist of the birds of the world. Version 7. Available at: https://datazone.birdlife.org/userfiles/file/Species/Taxonomy/HBW-BirdLife_Checklist_v7_Dec22.zip, referenced as BLI 2022.
Where the treatment varies between these sources, it may be noted by comments in the text or by presenting each variation with comments. Eg. subspecies cooki of Red-fronted Parakeet Cyanoramphus novaezelandiae in WLAB 4.1 is elevated to species level (Norfolk Island Parakeet Cyanoramphus cookii) in NZCL 2022. See Taxonomic changes since HANZAB book published in Red-crowned Parakeet (the NZCL 2022 name for Red-fronted Parakeet) or HANZAB Index+ for the treatment adopted.
[Information from Volumes 1, 3, 4 and 5,6,7]
From Vol. 1
The arrangement and nomenclature of orders, families and species closely follows those of Condon in the Checklist of the Birds of Australia, Part 1 (1975), and of Kinsky in the Annotated Checklist of the Birds of New Zealand (1970), both of which are essentially those of Peters’ Checklist of Birds of the World, Volume 1 (1979), and that in itself is much the same as the Wetmore order of 1960. At the generic level, admitted to be the most arbitrary, controversial and mutable taxon in the system, we have avoided sub-genera for the most part and thus differ from the above standards in one way or another. In some genera (Pterodroma, Sula and Phalacrocorax), we do so because proposed sub-genera seem not yet to be widely accepted; in Ardea, because the proposed sub-genera are hard or even impossible to separate. Further, we have treated some isolated, sedentary and insular forms of shags and ducks as full species rather than as subspecies because that seems to be more sensible biologically. Preparation of our first volume has coincided with the preparation of the Catalogue of Australian birds for the Australian Biological Resources Survey (ABRS) by Dr R. Schodde, and of the third edition of the New Zealand checklist. Originally we hoped that both would be published well before our first volume, but in the event we were obliged to commit ourselves taxonomically before they appeared. Thus, in spite of full cooperation and help from the compilers of these works and their helpers, we cannot be sure that our arrangement and those of the ABRS list and the new New Zealand checklist will be the same. Sibley, Ahlquist and Monroe (Auk 105: 409-23) have recently proposed a new classification of birds based on DNA-hybridization. Whether or not this comes to be generally accepted, it came too late for us to use. To have done so would have meant scrapping much work and preparation, and delaying production intolerably.
Names of the birds in Dutch, French, German, Japanese, Malay, Maori and Russian, as far as they are available, are listed in an appendix at the end of the book. There is also a separate appendix for Aboriginal names. The official attitude of the RAOU to the use of English names is set out in the Supplement to Emu 1977, Vol. 77 (Recommended English Names for Australian Birds). It favours an international rather than a parochial or insular approach to the matter and we have done so throughout. We have departed from the Recommended English Names in two cases: we have used Salvin’s Prion for Pachyptila salvini (in place of Lesser Broad-billed Prion; see explanation in masthead of P. salvini) and Australian White Ibis (in place of Sacred Ibis) for Threskiornis molucca, which we have considered a separate species. English names for species endemic to New Zealand are usually taken from the Annotated Checklist of the Birds of New Zealand (Kinsky 1970) and amendments (Notornis 27, Suppl.).
From Vol. 3
NOMENCLATURE
The taxonomy, nomenclature and arrangement of orders, families and species in Volume 3 follow those of the revised species list of Australian birds by Christidis & Boles (1994), except for the position Of the Scolopacidae within the Charadriiformes. This exception became unavoidable with the split of the Charadriiformes between Volumes 2 and 3, and because work on Volume 2 was too advanced to follow Christidis & Boles (1994) fully, Following Christidis & Boles (1994), the sequence of families Of Charadriiformes that occur in the HANZAB region is: Pedionomidae, Scolopacidae, Rostratulidae, Jacanidae, Chionididae, Burhinidae, Haematopodidae, Recurvirostridae, Charadriidae, Glareolidae and Laridae. In Our sequence, the Scolopacidae are out of place.
In this Volume, the taxonomic ranking given to the groups that comprise the Laridae follows Christidis & Boles (1994), but differs from that given in the introduction to the Charadriiformes in HANZAB 2.
English names for birds of Australia and Australian territories follow those ofChristidis & Boles ( 1994); English names for species endemic to New Zealand follow those of NZCL (1990).
Botanical names In the Laridae and Columbidae, the names of plants in all sections other than Food were checked by G. Carr using the following authorities:
- Allan, H.L. 1961. Flora of New Zealand. 1. R.E. Owen, Gov’t Printer, Wellington.
- Australian Biological Resources Study. 1993. Flora Of Australia. 50. Oceanic Islands. AGPS, Canberra.
- Hnatiuk, R.J. 1990. Census of Australian Vascular Plants. Aust. Flora & Fauna Ser. II; Bureau Flora & Fauna. AGI’S, Canberra. Moore, L.B., & E. Edgar. 1970. Flora of New Zealand. 11. A.R, Shearer, Gov’t Printer, Wellington.
- Wallis, J.C., & H.R. Airy Shaw. 1973. A Dictionary of the Flowering plants and Ferns. Cambridge Univ. press, Cambridge.
From Vol. 4
TAXONOMY AND NOMENCLATURE
As in Volume 3, we continue to follow the arrangement and nomenclature of Christidis & Boles (1994) and amendments (Christidis & Boles In press). For details of subspecies and subspecific nomenclature, we have followed Schodde & Mason (1997) where it was available except in cases where it conflicted with species limits set out in Christidis & Boles (1994, In press). Any deviations from the taxonomy of Schodde & Mason (1997) are explained within the texts. The arrangements of the few species recorded in the wider HANZAB region that were not included within these publications were determined in consultation with L. Christidis & W.E. Boles, as members of Birds Australia’s Taxonomic Advisory Committee, based on the principles and sources used by them in compiling their 1994 publication. English names follow those of Christidis & Boles (1994); English names for species endemic to NZ follow those of OSNZ (1990).
REFERENCES
- Christidis, L., & W.E. Boles. 1994. The Taxonomy and Species of the Birds of Australia. RAOU Monogr. 2. RAOU, Melbourne.
- —, — In press. Emu.
- OSNZ (Ornithological Society of New Zealand) . 1990. Annotated Checklist of New Zealand Birds and the Ross Dependency, Antarctica. Third edn. Random Century, Auckland.
- Schodde, R., & I.J. Mason. 1997. Zoological Catalogue of Australia. 37.2. Aves. CSIRO Publ., Melbourne.
From Vol. 5, 6 and 7 (repeated in each volume)
TAXONOMY AND NOMENCLATURE
Birds
The publication of the ground-breaking Directory of Australian Birds: Passerines by Schodde & Mason (2000 [abbreviated throughout this volume as DAB]) has provided a substantial base from which to further investigate the variation in species and, of greater import, the subspecies of Aust. birds, and has greatly assisted us in the preparation of this volume. For the first time in contemporary ornithology in the Aust. region, DAB presents a complete listing and analysis of the terminal taxa of Aust. passerine birds.
As in Volumes 3 and 4, for families and species we continue to follow the arrangement and nomenclature of Christidis & Boles (1994) and amendments (Christidis & Boles In prep.); the latter will incorporate several changes published in DAB. The departures from Christidis & Boles (1994) recognized in this volume are: separation of Short-tailed Grasswren Amytornis merrotsyi from Striated Grasswren A. striatus; recognition of Kalkadoon Grasswren Amytornis ballarae as a species separate from Dusky Grasswren A. purnelli; and separation of Western Wattlebird Anthochaera lunulata from Little Wattlebird A. chrysoptera. In this and subsequent volumes, details of subspecies and subspecific nomenclature essentially follow DAB except in cases where it conflicts with species limits set out in Christidis & Boles (1994, In prep.). However, even in those instances, subspecific treatment of DAB is always discussed within the texts and reasons for departure from DAB are given.
The arrangements of the few species recorded in the wider HANZAB region that were not included within the above publications were determined in consultation with L. Christidis and W.E. Boles (representing Birds Australia’s Taxonomic Advisory Committee and a member of the HANZAB Steering Committee), based on the principles and sources used by Christidis & Boles in compiling their 1994 publication. For NZ species, scientific nomenclature follows OSNZ (1990) except in cases where it conflicts with Christidis & Boles (1994, In prep.).
English names follow those of Christidis & Boles (1994); English names for species endemic to NZ follow those of OSNZ (1990).
Plants and animals other than birds
All scientific names, other than those of birds, were checked against the following references; for those groups for which volumes have been published, we have used the multi-volume series the Flora of Australia, the Fauna of Australia and the Zoological Catalogue of Australia.
Plants
For Aust., Hnatiuk (1990), ABRS (1993) and, for specific families, George (1986, 1989), Chippendale (1988) and Orchard (1995, 1998); for NZ, Allan (1961), Poole & Adams (1963), and Moore & Edgar (1970); and, more generally or outside these areas, Wallis & Airy Shaw (1973). We have retained Eucalyptus as a single genus, though we have often placed the subgeneric name Corymbia in brackets after Eucalyptus for species of bloodwoods.
Animals
- GENERAL INVERTEBRATES: Marshall & Williams ( 1972).
- MOLLUSCS: Vaught (1989) .
- SPIDERS: Main et al. (1985).
- INSECTS: Taylor et al. (1985); Lawrence et al. (1987), Campbell et al. (1988), Common (1990), CSIRO (1991), Naumann (1993), Lawrence & Britton (1994) and Nielsen et al. (1996) .
- FISH: Paxton et al. (1989) , Eschmeyer (1990) and Gommon et al. (1994).
- AMPHIBIANS AND REPTILES: Cogger et al. (1983 ) and Cogger (1992).
- MAMMALS: Bannister et al. (1988) and Strahan (1995).
REFERENCES
- Allan, H.L. 1961. Flora of New Zealand. I. R.E. Owen, Govt Printer, Wellington.
- ABRS (Aust. Biol. Resources Study). 1993. Flora of Australia. 50. Oceanic Islands. Aust. Govt Pub!. Service, Canberra.
- Bannister, J .L. , et al. 1988. Fauna of Australia. 5. Mammalia. Aust. Govt Publ. Service, Canberra.
- Campbell, I., et al. 1988. Zoological Catalogue of Australia. 6. Ephemeroptera, Megaloptera, Odonata, Plecoptera, Trichoptera. Aust. Govt Publ. Service, Canberra.
- Chippendale, G.M. 1988. Flora of Australia. 19. Eucalyptus, Angophora (Myrtaceae). Aust. Govt Publ. Service, Canberra.
- Christidis, L., & W.E. Boles. 1994. RAOU Monogr. 2.
- Cogger, H.G. 1992. Reptiles and Amphibians of Australia. Rev. edn. Reed, Sydney.
- —, et al. 1983 . Zoological Catalogue of Australia. 1. Amphibia and Reptilia. Aust. Govt Publ. Service, Canberra.
- Common, I.F.B. 1990. Moths of Australia. Melb. Univ. Press, Melbourne.
- CSIRO. 1991. Insects of Australia. CSIRO Publ., Melbourne.
- Eschmeyer, W.N. 1990. Catalogue of the Genera of Recent Fishes. Calif. Acad. Sci. , San Francisco.
- George, A.S. (Ed.) 1986. Flora of Australia. 46. Iridaceae to Dioscoreaceae. Aust. Govt Publ. Service, Canberra.
- — 1989. Flora of Australia. 3. Hamamelidales to Casuarinales. Aust. Govt Publ. Service, Canberra.
- Gommon, M.F. , et al. (Eds) 1994. The Fishes of Australia’s South Coast. State Printer, Adelaide.
- Hnatiuk, R.J. 1990. Census of Australian Vascular Plants. Aust. Flora & Fauna Ser. 11 (Bureau Flora & Fauna): Aust. Govt Publ. Service, Canberra.
- Lawrence, J.F. , & E.B. Britton. 1994. Australian Beetles. Melb. Univ. Press, Melbourne.
- —, et al. 1987. Zoological Catalogue of Australia. 4. Coleoptera. Aust. Govt Publ. Service, Canberra.
- Main, B.Y., et al. 1985. Zoological Catalogue of Australia. 3. Arachnida. Aust. Govt Publ. Service, Canberra.
- Marshall, A.J. , & W.O. Williams. 1972. Textbook of Zoology: Invertebrates. Macmillan, Melbourne.
- Moore, L.B., & E. Edgar. 1970. Flora of New Zealand. II . A.R. Shearer, Govt Printer, Wellington.
- Naumann, I. 1993. CSIRO Handbook of Australian Insect Names. CSIRO Publ., Melbourne.
- Nielsen, E.S., et al. 1996. Checklist of the Lepidoptera of Australia. Monographs on Australian Lepidoptera, Vol. 4. CSIRO Publ., Melbourne.
- Orchard , A.E. (Ed.) 1995. Flora of Australia. 16. Elaeagnaceae, Proteaceae 1 . CSIRO Publ., Melbourne.
- — 1998. Flora of Australia. 12. Mimosaceae (excluding Acacia), Caesalpiniaceae. CSIRO Publ., Melbourne.
- OSNZ (Orn. Soc. of NZ [E.G. Turbon, Convenor, Checklist Committee]). Checklist of the Birds of New Zealand and the Ross Dependency , Antarctica. Third edn. Random Century, Auckland.
- Paxton, J.R., et al. 1989. Zoological Catalogue of Australia. 7. Pisces. Aust. Govt Publ. Service, Canberra.
- Poole, A.L., & N.M. Adams. 1963 . Trees and Shrubs of New Zealand. R.E. Owen, Govt Printers, Wellington.
- Schodde, R., & I.J. Mason. 2000. Directory of Australian Birds: Passerines. CSIRO Publ., Melbourne.
- Strahan, R. (Ed. ) 1995. The Mammals of Australia. Reed New Holland, Sydney.
- Taylor, R.W., et al. 1985 . Zoological Catalogue of Australia. 2. Hymenoptera. Aust. Govt Publ. Service, Canberra.
- Vaught, K.C. 1989. A Classification of the Living Mollusca. Am. Malacologist Inc., Melbourne, Florida.
- Wallis, J. C., & H.K. Airy Shaw. 1973 . A Dictionary of the Flowering Plants and Ferns. Cambridge Univ. Press, Cambridge.
TREATMENT AND PRESENTATION
[Information from Volumes 1 and 5]
From Vol. 1
The bulk of the book is set out in standard systematic form with brief introductory remarks for taxa above the level of genus, written from the point of view of Australia, New Zealand and Antarctica, and detailed accounts for each species. The introductions for orders characterize the sorts of birds concerned, list constituent families and outline taxonomic arrangements that differ from ours. This may be transferred to the introduction for a family where there is only one family in the order. Introductions for families usually cover the types of birds concerned, number of sub-taxa and informal groupings such as superspecies, world distribution and representation in our region, and chief morphological and behavioural characters. Below the level of species we have avoided as far as possible any treatment of subspecies except in the paragraph for Geographical Variation at the end of the Plumages section. Subspecific discrimination is a valuable and necessary tool in museum studies and with birds in the hand, but not generally in the field.
The species accounts are divided into sections for Field Identification, Habitat, Distribution, Movements, Food, Social Organization and Behaviour, Voice, Breeding, and Plumages and related matters. If information on population is available it is included in the section on Distribution. The original plan was to have an editor for each section and to solicit an account for each species from experts, which the editors would then treat for consistency of style, arrangement and presentation. This simply did not work. For various rear sons, some of those who originally agreed to be editors could not carry on through the long period of preparation, and many contributors could not meet our onerous demands. Many ad hoc reorganizations had to be made and so we cannot give a simple list of editors and contributors. Detailed acknowledgements are made below. Details of the scope of each section, with an explanation of conventions and abbreviations used and special problems, are discussed in separate introductions for each section below. Some abbreviations and conventions are used throughout the work; others apply only to a particular section. The general ones are explained separately (see Abbreviations and Conventions).
About 900 species of birds have been recorded within our limits, depending somewhat on the classification used and including vagrants, introduced species with viable feral populations, and those extinct within historical times. In this volume we cover 196 species, of which most (162) breed within our limits and so receive full treatment in ten sections, as mentioned above. Many pelagic and Antarctic birds are, of course, rare and seldom found in Australian and New Zealand waters but breed in Antarctica or on subantarctic islands. Others (27) are regarded as non-breeding visitors, vagrants or accidentals, though some could be more regular migrants annually to Australian or New Zealand seas or to little-known areas on land. For them, sections on Food, Social Organization and Behaviour, Voice and Breeding are omitted, as may be detailed descriptions of plumage if material for study was not available (however, some very recent occurrences or vagrants to Antarctica or subantarctic islands have only a short paragraph). All these breeding and non-breeding species and most vagrant species are illustrated in colour, to show the plumages that can be identified in the field, from downy young to breeding adult. Further, there are seven species that are extinct in historical times or that have not been recorded since 1900 or for which we think that the record is doubtful, definitely erroneous or otherwise unacceptable. These are not illustrated and have only a short paragraph, setting out the evidence. Fossil and subfossil species are not covered.
From Vol. 5
The bulk of the book is set out in standard systematic form with brief introductory remarks for taxa above the level of genus, written from the point of view of Australia, New Zealand and Antarctica (see description of the HANZAB region below), and detailed accounts for each species. The only order introduction in this and subsequent volumes, for the Passeriformes, characterizes the sorts of birds within the Order, lists the constituent families in the HANZAB region and outlines taxonomic arrangements that differ from ours. A few morphological and behavioural characters common or frequent in the Order are also summarized. Introductions for families usually cover the types of birds concerned, number of sub-taxa and informal groupings such as superspecies, world distribution and representation in our region, and chief morphological and behavioural characters common or frequent in the family. Some aspects of behaviour, such as resting postures, comfort behaviour, including head-scratching, bathing and preening, and thermoregulation are characteristic of whole families rather than of individual genera or species; they are discussed or mentioned here rather than in the species accounts where they do not fit in easily and would need to be repeated time and again.
The species accounts are divided into sections for Field Identification, Habitat, Distribution and Population, Threats and Human Interactions, Movements, Food, Social Organization, Social Behaviour, Voice, Breeding, and Plumages, Bare Parts, Moults, Measurements, Weights, Structure and Geographical Variation; in some circumstances, additional sections for Ageing, Sexing and Recognition follow Structure. Each account concludes with a fu ll list of references. Throughout the work, detailed descriptions and summaries are largely confined to data collected in the HANZAB region (see below) and extralimital data are not usually presented in any detail, though important useful references are cited . We do, however, present a little more detail from New Guinea sources where there is little or no information for Australia or New Zealand. Details of the scope of each section, with an explanation of conventions and abbreviations used and problems specific to those sections, were discussed fully in introductions for each section in Volume 1 and revised introductions for most sections appear below.
Breeding species receive full treatment in these sections. For non-breeding migrants, the sections on Social Organization, Social Behaviour and Breeding are omitted. For species that are accidental to the HANZAB region , only the sections on Field Identification, Habitat, Distribution, Movements and Plumages and related matters are covered. In these categories, some species, such as introduced species, may have a reduced treatment for Plumages and related matters if there is an adequate summary published elsewhere (see the introduction to Plumages and related matters). For species that have become extinct since European settlement, we summarize as much as we know of the biology of the species, and prepare as full an account for Plumages and related matters as possible with the material available in museums or elsewhere. However, for many extinct species, there is little or no information on the biology of the birds and few specimens available to us, so we have been able to do little.
Lastly, some species receive only a brief treatment, with a summary paragraph outlining the occurrence or claimed occurrence in the HANZAB region. Such species include: vagrants to the wider region covered in HANZAB but beyond the generally recognized limits of Australia and New Zealand and their territories (for example, in this volume, two species vagrant to South Georgia from the Americas ); unverified reports or claims for the region; and failed introductions to the HANZAB region. Where there are many failed introductions within a family, they may all be dealt with together.
Some abbreviations and conventions are used throughout the work; others are applied only to a particular section. All abbreviations and conventions are listed [see Abbreviations and Conventions]. The rest of this introduction largely uses those abbreviations and conventions.
THE HANZAB REGION
The region covered by HANZAB is: Aust. within the limits of the Continental Shelf, including the reefs and islands of the Coral Sea, N to 10 S or the Qld-New Guinea political border, whichever lies farther N, but excluding the e. end of New Guinea and adjacent islands above 10 S; the Aust. external territories of Cocos-Keeling, Christmas (in the Indian Ocean)l, Lord Howe, Norfolk, Heard and Macquarie Is; NZ and its islands, from the Kermadec Grp in the N to Campbell I. in the S and the Chatham Grp in the E; the Antarctic Continent; and the subantarctic islands, ncluding Marion, Prince Edward, Iles Crozet, Iles Kerguelen, the islands of the Scotia Arc: South Georgia, South Sandwich, South Orkney and South Shetland Is, and the subantarctic territories of Aust. and NZ already mentioned. The boundaries of the region are shown on the various maps. [see Gazetteer]
FIELD IDENTIFICATION
[Information from Volumes 1, 3 and 4]
From Vol. 1
This section sets out the characters by which a species may be identified in the field, even without the help of illustrations. It goes without saying that the recognition of many features often or usually depends on the circumstances and wear of plumage, especially for birds at sea, and this should be kept in mind when making and evaluating observations. For the most part we have not presented differences between subspecies here, because we do not wish to encourage the idea that sub-species can be identified in the field, except in the case of a few, very well-marked examples. References are not usually given in this section.
[Note the mention of paragraphs here may not be appropriate as some may have been broken into multiple paragraphs in the online version.]
The presentation is in four paragraphs. The first is designed to help those who are ignorant of A’asian birds, or even of birds in general, to decide whether they are on the right track for identification. It gives a rough indication of the size, shape, appearance and type of bird being described. Measurements of total length, wingspan and weight are given in gross terms, as a guide to the size of the bird, and, where necessary with a broad indication of the proportions of head and neck, body and tail (detailed measurements are in the Plumages section). Brief mention is then made of outstanding characters of plumage or other features, especially if they are diagnostic, and it is indicated whether differences occur by age, season or sex.
The second paragraph (DESCRIPTION) describes the various stages of plumages as seen in the field. Here, as necessary, descriptions are given of adult male and female, breeding and non-breeding, downy young, juveniles and immature stages, as well as morphs and phases. After the first mention of a character, it is not usually repeated, only the differences being emphasized. It is hoped that with this information on the different stages in plumage, field observers will be encouraged to discriminate more carefully between sexes and ages than is often done, because important data on patterns of movement, age at first breeding and social behaviour can thus be collected.
The third paragraph (SIMILAR SPECIES) sets out those species that may be confused with the species in question, sometimes even to a point that may seem ridiculous or impossible. Field conditions, however, can play some strange tricks and one must be careful. There is nothing worse than publishing a doubtful or incorrect identification as a certainty, which will be perpetuated and is hard to eradicate or correct (see discussion in Distribution introduction). Some care has been taken to make comparisons between species in the same way: species X, Y or Z always first, smaller, paler, etc, than species A, which is the subject of the account.
The fourth paragraph tries to give an outline of less concrete aspects of identification and is generally the weakest part of the section. It is not easy to remember to record aspects of habitat, gait, swimming, flight and so on, which one assumes are perfectly obvious and well known. There is probably much scope for improvement here, because contributors and editors had a good deal of trouble in covering the field, even as a general outline. Usually fuller information (and references) on various aspects discussed here may be found in other sections. It has been thought necessary to condense substantially the very detailed descriptions and comparisons submitted by some contributors for the second and third paragraphs. It is hoped that such condensation has not gone too far and that nothing really important has been left out.
From Vol. 3
FIELD IDENTIFICATION
The approach to this section remains much as outlined in the Introduction to Volume 1, though several aspects deserve explanation or comment.
For paragraph one, it was difficult to obtain accurate values for length and wingspan and to establish relative size within genera or families. Published data vary greatly and, in some cases, we had considerable doubts about the accuracy of published data (e.g. the wingspans for many Charadriiformes given in Volumes 3 and 4 of BWP appear consistently too high). Whenever good data from museum specimens were available for birds collected in the HANZAB region, we used these in preference to published data. Otherwise, data were taken mainly from the following sources. WADERS: BWP (Volumes 3, 4); Hayman et al. (1986); Chandler (1989) and Paulson (1993); SKUAS, JAEGERS, GULLS AND TERNS: mainly Harrison (1983, 1987), BWP (Volumes 3, 4), and Olsen & Larsson (1995); COLUMBIFORMES: Frith (1982), Crome & Shields (1992), Pizzey (1980) and Slater et al. (1989). For relative size within genera or families, we relied entirely on information in the foregoing literature; Paulson (1993) proved particularly useful for many of the waders, as did Olsen & Larsson (1995) for the terns. In light of the difficulties encountered during preparation of this and previous volumes, we strongly encourage museum workers and others to help obtain and make available accurate measurements of length, wingspan and weight for A’asian birds, which would be invaluable for use in
future volumes of HANZAB.
Species accounts
The Charadriiformes present considerable difficulties in identification and ageing in the field. Fortunately, recent years have seen the appearance of a wealth of detailed specialist identification papers and guides covering waders, gulls and other groups, and many field identification problems previously considered almost impossible to resolve (e.g. separation of stints in juvenile plumage) are now possible
or even routine in some instances. Resolution of these difficult identification problems has come about largely through adoption of the so-called ‘new approach’ to identification (see Grant & Mullarney 1989), with its emphasis on topography and moult of birds as well as traditional skills of bird identification. With widespread use of telescopes and specialist identification guides, birdwatchers are nowadays scrutinising birds more closely than ever before and in much greater detail, as they attempt not only to identify a bird to species but also to determine its age, sex and stage of moult where possible. We have attempted to summarize all characters important in identification, ageing and sexing. The sections of Plumages and related matters are complementary to the Field Identification section and need to be consulted for more detailed information on patterns of individual feathers and of moult. An exception is made for those few very rare vagrant species where only a brief Plumages account is given (usually only when extralimital summaries are already available, e.g. in BWP); in these cases, the Field Identification accounts are more detailed than is normally the case. We have occasionally provided references for some particularly difficult identification problems. For a full review of the new identification techniques, plumages, topography, judgement of size and structure and other aspects of the new approach, see Grant & Mullarney (1989).
GLOSSARY
Some terms have been introduced into the accounts or have been used again but were not previously defined.
- APICAL SPOT: white tips of primaries of gulls.
- CARPAL BAR: band of dark feathers extending diagonally across the inner upperwing, from the carpal joint to the base of the tertials, and contrasting with paler rest of wing; formed by median secondary coverts and rear rows of lesser secondary coverts. Characteristic of many gulls.
- COMMIC TERNS: a group of very similar medium-sized Sterna terns: Roseate S. dougallii, White-fronted S. striata, Common S. hirundo, Arctic S. paradisaea, Antarctic S. vittata and Kerguelen S. virgata.
- CUBITAL BAR: band of dark feathers along the leading-edge of the inner upperwing, and contrasting with paler rest of wing; formed by lesser secondary coverts. Occurs in many terns.
- HOOKBACKS: dark markings on the outer primaries of some terns, in which dark areas on tips of the outer webs extends on to the inner webs as a dark line along inner edge; see illustrations Fig. 8, Antarctic Tern.
- INNERWING-COVERTS: Secondary coverts. In this volume, used mainly to refer to those coverts visible on the folded wing of a standing bird.
- INNERWING: secondaries and secondary coverts (including tertials and their coverts).
- LINING OR WING-LINING: primary and secondary coverts of underwing.
- MOULT-CONTRAST: an obvious difference in colour and wear between adjacent feathers of different ages. A classic example occurs in adult breeding Common Terns, in which the contrast between newer paler inner primaries and older darker and more worn outer primaries on the upperwing forms a diagnostic field character.
- OUTERWING: primaries, primary coverts and alula.
- PRIMARY PROJECTION: on a folded wing, the distance primaries project beyond the longest tertial compared with the length of the exposed tertials.
- SADDLE: the mantle, back and scapulars together.
- SCAPULAR CRESCENT: narrow pale crescent formed by white tips of rearmost scapulars, often prominent on standing gull or tern.
- SECONDARY BAR: contrasting dark band on inner upperwing, formed by dark bases of secondaries.
- TAIL-STREAMERS: specialised rectrices (usually long and pointed) that project beyond other rectrices. Examples in this volume are tl of adult breeding jaegers and t6 of many terns.
- TERTIAL CRESCENT: narrow to broad pale crescent formed by white tips of longest tertials, often prominent on standing gull or tern.
- UNDERBODY: ventral body plumage, not including underwing and undertail.
- WING-POINT: in the Field Identification accounts, refers to that part of the wing-tip visible beyond the longest tertial on a folded wing; see also primary projection. For birds in the hand, refers to the longest primary on the folded wing.
REFERENCES
- Chandler, R.J . 1989. North Atlantic Shorebirds. MacMillan, Lond.
- Crome, F.J.H., & J. Shields. 1992. Parrots and Pigeons of Australia.
- Angus & Robertson, Sydney.
- Frith, H.J. 1982. Pigeons and Doves of Australia. Rigby, Sydney.
- Grant, P., & K. Mullarney. 1989. The New Approach to Identification. Peter Grant, Ashford, Kent, UK.
- Hayman, P., et al. 1986. Shorebirds. Croom Helm, Sydney.
- Harrison, P. 1983. Seabirds: An Identification Guide. Croom Helm,
- Lond.
- — 1987 Seabirds of the World: A Photographic Guide. Christopher
- Helm, Lond.
- Olsen, K.M., & H. Larsson. 1995. Terns of Europe and North
- America. Christopher Helm, Lond.
- Paulson, D. 1993. Shorebirds of the Pacific Northwest. Univ. Washington,
- Seattle.
- Pizzey, G. 1980. A Field Guide to the Birds of Australia. Collins,
- Sydney.
- Slater, P., et al. 1989. The Slater Field Guide to Australian Birds.
- Lansdowne Press, Sydney.
From Vol. 4
FIELD IDENTIFICATION
Generally, this section remains much as described in the Introduction to Volume 3, though we have changed the arrangement a little to try to remove further duplication between Field Identification and the sections of Plumages and related matters. The latter sections complement Field Identification and need to be consulted for detail on patterns of individual feathers and of moult. The first paragraph has been expanded to include the descriptions of field characters important in identification, ageing and sexing, concentrating on the overall appearance of the birds. The discussion of similar species also changes focus, from presenting details of the similar species to presenting those of the species under consideration. At times, where finer detail than is provided in the preceding description is required to distinguish similar species, such detail is usually given in the discussion of similar species, where comparisons can be directly made with the characters of the similar species.
HABITAT
[Information from Volumes 1 and 5]
From Vol. 1
The ideal habitat description for a particular species of bird presents an analysis of the critical factors determining distribution and the suitability of particular sites, taking into account needs for different purposes (e.g. feeding, breeding, roosting, moult); of the dynamics of use of habitat daily, seasonally or for longer periods; and of the effects of alteration of habitat, naturally or by human agency. It ought to apply throughout the species’ geographical range, or, at least, a large part of it. It ought to be predictive as well as descriptive and present possibilities and ideas for management.
By these criteria, there is scarcely a species within our region for which a comprehensive description of habitat can be compiled. The difficulties are the same as encountered in BWP and apply globally; particularly the lack of detailed studies for most species and the imprecise and inconsistent use of terms of description.
The most intractable difficulty is the lack of comprehensive studies on use of habitat by birds. This is a problem worldwide, but is exacerbated for us by low density of population, remoteness, inaccessibility and severe climatic conditions in some parts. In compiling these texts, the authors and editors have attempted to integrate and condense published information to produce a generalization for each species. For some species, we were fortunate to be able to draw on systematic studies. Also valuable were the observations and insights of experienced field observers. But all too often, in the absence of these sources, we gleaned information from numbers of short notes, papers on other topics, and annotated bird lists, much of it inevitably anecdotal, superficial and fragmentary. Even where there have been studies of habitat, often the focus is on aspects that are obvious or most amenable to study; for example, there is little quantitative information on use of airspace and underwater zones, and the study of the marine ecology of seabirds in our region is still in its infancy. Information from small areas is rarely interpreted with a view to integrating it into the wider geographical picture, and, perhaps most frustratingly, much valuable information remains unpublished or exists only in sources that are difficult to find.
Thus far, studies of avian habitats in our region have generated a bewildering variety of approaches to classification of habitat. The criteria used have in general been subjectively chosen, without evidence that they are relevant in determining the occurrence and abundance of birds, and many are appropriate only over small areas. Even where there are systems of classification covering wide areas and in widespread use in other disciplines, they have as yet received little attention from ornithologists e.g. Specht’s (1981) structural classification of vegetational formations in Aust.
In compiling this section, therefore, we have used terms for habitat description that are familiar and regularly used by both amateur and professional ornithologists, assigning to them reasonably precise meanings. Any more restrictive approach would exclude most information available at present. A glossary follows This presents terms used in the text that may be unfamiliar to some readers or that have other meanings in common usage in our region.
The Habitat section for each species discusses in sequence the biogeographical settings of distribution, broad terrestrial and aquatic groupings, details of habitats used for foraging, breeding, roosting and moulting, and relations with humans where these are pertinent. Although it is difficult to choose references for citation in a work such as this, we have attempted to provide primary sources, especially acknowledging sources of significant facts and studies of particular species or groups. It is impossible that such listings should be exhaustive. This work holds, with few exceptions, the first attempts to integrate all information available on habitat use by bird species using the A’asian and Antarctic regions. Probably the work’s most important function will be to stimulate further studies and encourage publication of existing information.
GLOSSARY
This glossary defines the principal terms used for habitat description in our text, in recognition of the need to standardize and increase the precision of such terms. A number of other terms are used throughout according to general English usage and are not defined here. References used in producing this compilation are Moore (1949), Press & Siever (1978), Corrick & Norman (1980), Gosper (1981), Pearce (1981), Specht (1981), Corrick (1982), Ainley & Boekelheide (1983), Ainley et al. (1984), McDonald et al. (1984), Aust. Atlas, and BWP.
- ACACIA SCRUB. Vegetation dominated by shrubs of the genus Acacia; includes open-scrub, tall shrubland, tall open-shrubland and low open-shrubland of Specht (1981).
- ANABRANCH (anastomosing plus branch). Branch that leaves river and re-enters it downstream.
- ANTARCTIC CONVERGENCE. See Polar Front.
- ANTARCTIC SLOPE FRONT. Oceanic zone overlying Antarctic continental slope, where shelf-water meets circumpolar deep water, and strong gradients of temperature, salinity and turbidity occur.
- AQUATIC VEGETATION. Plants growing in water; may reach but not project above surface.
- ARID ZONE. Regions where mean annual rainfall is less than 250 mm.
- ATOLL. Coral reef in the shape of a ring or horseshoe, broken or continuous; enclosing a lagoon.
- BACKWASH. Return flow of water down beach after wave has broken.
- BILLABONG. Properly an ox-bow lake, formed when a meander of a river is cut off as the river modifies its course; popularly used for other water-bodies.
- BORE. Hole drilled in the ground from which underground water is pumped and reticulated.
- BOUNDARY CURRENTS. Fast-flowing currents concentrated along edges of major oceans. Poleward currents on western edges of oceans are very intense and are known as WESTERN BOUNDARY CURRENTS.
- BRAIDED RIVER (STREAM). Intricate system of interlacing channels, formed in wide river-beds choked with coarse sediments.
- CAY. Flat mound of sand built up on reef flat slightly above high-tide level.
- CLEAR-FELLING. Forestry operation in which all trees on a site are cut down.
- CLIMATIC ZONES. GLOBAL. Five main zones into which Earth is divided according to climate. Comprise Tropical Zone: region lying between the Tropics of Cancer (23°27’N) and Capricorn (23°27’S); Frigid Zones: regions enclosed by Antarctic Circle (66°33’S) and Arctic Circle (66°33’N); Temperate Zones: regions lying between Tropical and Frigid Zones. MARINE. Climatic zones of oceanic surface water defined by Ainley & Boekelheide (1983). Tropical Zone: waters with sea surface-temperature (SST) of at least 22.0 °C. Subtropical Zone: SST 14.0-21.9 °C. Subantarctic Zone: SST 4.0—13.9 °C. Antarctic Zone: SST below 4.0 °C.
- CONTINENTAL SHELF. Underwater plateau extending from coast to a depth of about 200 m; shelf-waters: zone of water over the continental shelf.
- CONTINENTAL SLOPE. Beyond edge of continental shelf, ocean floor slopes to the abyssal plain (often at depths >4000 m). Worldwide, slope averages 4° but round Aust. may be up to 40° (Bunt 1987).
- CREEK. Stream of less volume than a river; small tidal channel through a coastal marsh; wide arm of a river or bay. Popularly applied in Aust. to any, rather small, drainage channel or waterway, permanent or impermanent, inland or coastal.
- DAM. Small (<10 ha), artificial water storage formed by excavation or impoundment; used for stock watering, irrigation or domestic supply in agricultural or pastoral regions.
- DRY SEASON. Season in monsoonal areas when little rain falls; usually Apr. to Nov. in ne. Aust.
- DUNE. Hill or ridge of sand formed by wind-blown sand or other granular material. CONSOLIDATED DUNE. Dune stabilized by cover of vegetation.
- EMERGENT VEGETATION. Plants projecting above canopy or water surface.
- EUTROPHICATION. Formation of superabundance of algal life in body of water, caused by influx of nutrients. FIORD. Former glacial valley with steep walls, now occupied by sea.
- FLOODPLAIN. Plain bordering a river; formed from sediments deposited during intermittent or seasonal flooding, and characterized by billabongs, swamps, meandering creeks.
- FOREST. Vegetation of trees, usually over 10 m high, with projective foliage cover of more than 50%; includes tall open-forest, open-forest and low open-forest of Specht (1981).
- FRONT (OCEANIC). Line or zone of separation at sea surface between water-masses of different physical characteristics, particularly temperature.
- GIBBER PLAIN. Level land covered with pebbles, usually in arid regions; little vegetation; barren stony waste.
- GUANO. Compacted mass of faeces of colonial species of birds; accumulated over many years.
- HEATH. Vegetation dominated by shrubs; includes closed-heathland, open-heathland and dwarf open-heathland of Specht (1981).
- HERB. Non-woody plant.
- ICE. Types discussed in the text are: SHELF-ICE: floating seaward extension of continental glaciers; SEA-ICE: ice formed by freezing of sea water; PACK-ICE: unattached sea-ice, varying from open to fully consolidated; FAST-ICE: sea-ice attached to shelf-ice or land; ICEBERG: mass of land-ice broken off from glacier and afloat at sea; ICE-FLOE: small mass of floating ice detached from pack-ice, limits usually within sight.
- IMPROVED PASTURE. Pasture to which fertilizer has been applied.
- ISLAND. Piece of land surrounded by water. Marine islands can be classified according to origin; CONTINENTAL ISLAND: formed by separation from continental mainland; OCEANIC ISLAND: formed in ocean independent of mainland; VOLCANIC ISLAND: volcanic in origin; CORAL ISLAND: built by action of coral polyps.
- ISOTHERM. Contour line joining points of equal temperature or equal average temperature; oceanic or atmospheric.
- KRILL. Marine crustaceans; Arthropoda, Crustacea, order Euphausiacea, Euphausia or Nyctiphanes. Form swarms in Antarctic and subantarctic seas.
- LAGOON. Strictly an enclosed coastal lake, pool or inlet, separated from ocean by broken or continuous banks of sand, earth or shingle; or waters enclosed by an atoll. In Aust. applied popularly to any rather shallow or small water-body such as billabong, pool or pond.
- LEVEE. Natural ridge along bank of creek or river formed by deposition of silt during flooding; also artificial barrier to floods constructed in similar form.
- LITTORAL. Intertidal area of sea or ocean.
- MALLEE. Multi-stemmed eucalypt growing from subterranean rhizome; also vegetation in which mallee is dominant; corresponds to open-scrub of Specht (1981).
- MANGROVE. Rhizophoraceae; many genera in Aust.
- MEADOW. Seasonal or transient shallow freshwater wetland characterized by cover of low emergent vegetation, particularly semi-aquatic herbs.
- MEANDER. Broad curves in creek or river forming as water erodes outer bank of curves and deposits sediment against inner bank.
- MONSOON. Climatic regime in which the wind blows in one direction for about half the year and in the opposite direction for the other half. Prominent in tropics on e. sides of continents; in ne. Aust., moist onshore winds prevail in summer.
- MONSOONAL REGIONS. Regions affected by the monsoon, and experiencing distinct wet and dry seasons. Within our limits, coastal and subcoastal ne. Aust. and adjacent islands.
- MORAINE. Deposit of debris and rock fragments at margin of glacier.
- PARK. Enclosed piece of public ground in urban areas, used for ornamental and recreational purposes; often planted with exotic grass, shrub and tree species, and containing artificial pools or lakes.
- POLAR FRONT. Circumpolar Zone where cold Antarctic surface-water sinks below less dense subantarctic surface-water; northernmost extent coincides with 2 °C subsurface isotherm.
- RAINFOREST. Dense forest growing in areas of heavy rainfall; trees are evergreen and predominantly broad-leaved; includes tall closed-forest, closed-forest, low closed-forest and closed-shrub of Specht (1981).
- REED. Herbaceous erect plant, particularly of the genus Phragmites.
- REEF. Ridge of rock or coral (CORAL REEF) in sea, just above or below the surface. RIP. Narrow, fast-flowing ocean current. RUSH. Herbaceous erect plant of the families Juncaceae, Typhaceae.
- SALT LAKE. Lake, usually in arid or semi-arid zone, where evaporation exceeds inflow, so that water highly saline; in arid Aust., usually dry with flat barren surface-deposit of salt.
- SALTBUSH. Vegetation in which chenopods are dominant, particularly Atriplex, Enchylaena, Rhagodia ; includes low-shrubland, low open-shrubland and very open sedgeland of Specht (1981).
- SALTFIELD. Set of ponds for production of salt by natural evaporation of seawater.
- SALTMARSH. Low-lying, flat land regularly or intermittently flooded by saline or brackish water and covered or fringed by halophytic vegetation; coastal or inland.
- SALTPAN. Semi-permanent saline wetland; some aquatic plants (e.g. Ruppia, Lepilaena) in shallow waters; little or no emergent vegetation.
- SCORIA. Congealed lava or lava fragments containing large number of vesicles.
- SCREE. See TALUS.
- SEAMOUNT. Submarine mountain rising at least 900 m above ocean floor.
- SEDGE. Herbaceous erect plant; Cyperaceae and some other families.
- SEMLARID ZONE. Regions with mean annual rainfall of 250–500 mm.
- SHRUB. Woody plant <8 m tall, with many branches and ample foliage; replaces BWP’s BUSH; in common usage in Aust. for remote or undeveloped country.
- SPINIFEX. Vegetational association in which mound-forming grasses, known collectively as spinifex, are dominant; Gramineae, Triodia and Plechtrachne.
- STACK. Rocky islet or pillar near coastline, isolated by erosive action of waves.
- SWAMP. Wetland area, permanent, seasonal or ephemeral; typically richly vegetated with emergent and aquatic plants. BWP classifies vegetated wetlands as MARSHES and SWAMPS on the basis of persistence of water, but the dry climate over much of our region ensures that few wetlands, shallow enough to support rich plant growth, are permanent.
- TALUS. Deposit of angular fragments of weathered rock accumulated at base of cliff or steep slope.
- TUSSOCK GRASSLAND. Grassland dominated by grasses forming discrete but open tussocks.
- UNDERSTOREY. Shrub or tree layer below uppermost stratum.
- VOLCANO. Vent in earth’s crust through which lava reaches surface; includes deposits surrounding vent.
- VOLCANIC ASH. Fine particles of lava ejected from volcano in eruption and deposited as sediment on land.
- VOLCANIC OR CINDER CONE. Conical hill built up of material ejected from volcano and deposited around outlet.
- WET SEASON. Season in monsoonal areas when most rain falls; usually Dec.—Mar. in ne. Aust.
- WOODLAND. Vegetation association of well-spaced trees less than 30m high; includes open-forest, low open-forest, woodland, low woodland, open-woodland and low open-woodland of Specht (1981).
REFERENCES
- Ainley, D.G., & R.J. Boekelheide. 1983. Studies avian Biol. 8: 2-23.
- Ainley, D.G., et al. 1984. AOU orn. Monogr. 32: 1-97.
- Bunt, J.S. 1987. Pp. 17-42. In Fauna of Australia. 1A.
- Corrick, A.H. 1982. Proc. R. Soc. Vict. 94: 69-87.
- Corrick, A.H., & F.I. Norman. 1980. Proc. R. Soc. Vict. 91: 1-15.
- Gosper, D.G. 1981. Corella 5: 1-18.
- Keast, A. (Ed) 1981. Ecological Biogeography of Australia.
- McDonald, R.C., et al. 1984. Australian Soil and Land Survey Field Handbook.
- Moore, W.G. 1949. A Dictionary of Geography.
- Pearce, A. 1981. CSIRO Div. Fish. Oceanogr. Rep. 132: 1-51.
- Press, F., & R. Siever. 1978. Earth.
- Specht, R.L. 1981. Pp 163-297. In: Keast 1981.
From Vol. 5
The problems in assembling the habitat texts were discussed in the introduction to Volume 1 and have proved to be common to all subsequent volumes. For nearly all of the species in the HANZAB region that we have dealt with so far, it has proved difficult, and sometimes impossible, to assemble even a general overview of use of habitat by birds, let alone an analysis of the critical variables of habitat for each species. There are few comprehensive studies on habitat use by birds in the region and the texts in HANZAB are, for most species, the first attempt at collating and synthesizing the diversity of information contained in a wide range of published and unpublished sources.
In addition to the lack of systematic study and analysis of habitat, the difficulty of assembling an overview of habitat use by birds in the HANZAB region is exacerbated by the lack of consistent, or even accurate, classification of habitats by both amateur and professional ornithologists. While this is understandable in early literature, it is disappointing to find this perpetuated in more recent systematic studies or annotated lists, especially given the availability of systems of habitat classification covering wide areas and in widespread use in other disciplines, e.g. Specht’s (1981) structural classification of vegetation formations in Aust. Happily, there are increasing numbers of studies that do adopt accepted systematic classifications of vegetation and other habitat variables.
Despite the problems just described, we have attempted to describe habitat with standard terms. For structural descriptions of rainforest and non-rainforest vegetation and descriptions of landforms, we have as far as possible used the definitions given in the Australian Soil and Land Survey Field Handbook (McDonald et al. 1984; also see AUSLIG 1990). Other terms we use commonly are given in the glossary associated with the introduction to Habitat in Volume 1 (and which is not repeated here). Equally, however, it is not always possible to convert the often vague and ill-defined descriptions found in the published literature (e.g. scrub) to standard terminology; where it is considered useful, we include such descriptions, usually without comment.
Given our experience in assembling habitat texts for this and previous volumes, particularly when dealing with the primarily terrestrial species of Volume 4 and the passerine volumes here and to come, we have changed the arrangement of this section. Previously, following the first paragraph, there were separate paragraphs that dealt with breeding, feeding and roosting and loafing habitats. These paragraphs have been abandoned in this and subsequent volumes because:
- such habitats are almost always the same as those already described in the preceding paragraphs or, occasionally, a subset of them; and
- because we usually have so few details of such habitats beyond some details of use of sites for these behaviours, and which are already summarized in Breeding (Site), Food (Behaviour) and Social Behaviour (Roosting).
Finally, the information concerning human interactions and modifications to habitat, formerly in the last paragraph, has been moved to a new section, Threats and Human Interactions (see below).
The usual arrangement in this and remaining volumes is as follows. Unless there is very little information in total, the section opens with a brief introductory paragraph that summarizes: the main habitat types used by the species; the biogeographical settings of distribution, identifying the climatic zones in which the species occurs; and, if there is information available, the commonly inhabited landforms in which the species occurs. General references are provided here, but the references provided in the following more detailed analysis must also be consulted for a complete listing.
The introductory paragraph is then followed by a synthesis of the available information on use of habitat by the species, dealing firstly with those habitats used most often or commonly, through to those used only infrequently. Use of modified habitats (such as urban areas or farmland) is also discussed, either separately at the end of the paragraph, or integrated with the main discussion, depending on the frequency of use of such habitats. Occasionally, information is presented separately for different subspecies or different biogeographical or climatic regions (e.g. for widespread species ). We have tried to cite all important primary sources, especially acknowledging sources of significant facts and studies of particular species or groups. It is impossible that such listings of references should be exhaustive. Lastly, where there are detailed studies, these are often now presented in a separate final paragraph and may include information on differential use of habitats in an area or region or details of studies of human impacts, such as fire or logging.
REFERENCES
- AUSLIG. 1990. Atlas of Australian Resources. 6. Vegetation. Third series. Aust. Surveying & Land Information Grp, Dept Administrative Services, Canberra.
- Keast, A. (Ed.) 1981. Ecological Biogeography of Australia. Junk, The Hague.
- McDonald, R.C., et al. 1984. Australian Soil and Land Survey Field Handbook. Inkata Press, Melbourne.
- Specht, R.L. 198 1. Pp 163-297 In: Keast 1981.
DISTRIBUTION AND POPULATION
[Information from Volumes 1, 2, 3, 5, 6 and 7
From Vol. 1
In this section we try to present a summary of the known distribution of each species, within our limits with a mention of extralimital range. Maps of distribution appear for all species except accidental vagrants and, like good cartoons, obviate the need for much text. However, they can be presented only on a small scale and so some fairly detailed explanation has to be appended to make them useful. Distribution is inextricably linked with movements though we try and avoid overlap with that section.
After a summary of world distribution of the species and its occurrence in our region, details of distribution are given, generally for Aust., NZ, the territories of Lord Howe, Norfolk, Christmas and Cocos-Keeling, Kermadec and Chatham Is, in that order, followed by records in subantarctic islands and vagrants elsewhere. For Antarctic and pelagic species, the main distribution is usually given first and then records in more temperate regions in the same order as above. Breeding distribution is then considered. The last paragraphs give estimates of population (if not already with breeding distribution), and the status of the species. The history of introductions and colonizations is outlined where necessary.
Breeding distribution and Population
It hardly needs saying that, as regards the speciation and classification of birds, a vital consideration is interbreeding with accompanying gene-flow. One assumes that this takes place only within the breeding range. It is on the basis of breeding range that the question of sympatry or allopatry is established. Thus, the breeding range of a species is surely what is fundamentally important from a biological point of view. However, there is surprisingly little information on the present breeding range of many A’asian species. Within Aust. it may be thought that the Atlas of Australian Birds and RAOU Nest Record Scheme would meet requirements, but the Atlas relied primarily on records of presence or absence, and evidence for breeding came a poor second. Records in the NRS are usually too scattered and fragmentary to be of much use. In consequence, one has no means of knowing whether breeding occurs in all blocks where the birds were seen during the Atlas work, though one may be quite certain that it did not do so. Later work, such as organized waterbird surveys in Vic. and WA, has enabled us to fill some gaps. For NZ, the situation is even worse because the Atlas of New Zealand Birds does not indicate breeding. For most colonially nesting species, each colony known to us is listed, with its size in recent years. However, for some species, such as ibises, only the larger or long-established colonies are listed, with references to sources. The estimated figures may or may not give a reasonable idea of populations at the present time. They ought, however, to provide a measure against which future fluctuations can be judged. They ought also to encourage people to fill the gaps that must be there. Similarly, figures of recent surveys of waterfowl are recorded, as an index of the numbers of birds seen rather than as actual censuses. All the same, there are no estimates of populations for many species.
Accidental and vagrant species. First records for species in Aust. and NZ, records of a species far outside its normal range and records of vagrants generally present difficulties. It is no use burking this question, and the fact is that, until recently, reporting and vetting of such records in Aust. has been deplorable. Until recently, there has been no official body to which records have to be submitted for critical appraisal and acceptance before publication. In consequence, publication has often been made without any acceptable supporting justification and acceptance has been lax. Indeed, we have found that some such records accepted in the Atlas of Australian Birds are not acceptable by modern standards.
In 1975 the RAOU established the Records Appraisal Committee for reviewing unusual records of all sorts. Unfortunately, for many reasons, the Committee has vetted few records and has not yet achieved much authority. The Committee has been reformed and is now evaluating published and unpublished records. At the same time, within the last 20 years or so, the standard of exact and critical observation in the field has been improved immensely by the enthusiasm and abilities of a new generation of field observers, particularly as regards waders and seabirds, and by the organizing of regular boat-trips across the continental shelf. In this way, a great deal of valuable information on occurrence, status and distribution of species hitherto little known in our region has accumulated, but unhappily not much of it has been published satisfactorily. This is fair neither to the observers, who have not received their due credit, nor to general ornithologists, who have to accept the observations on hearsay or not at all. In a work that is trying to assemble the facts, the situation is unsatisfactory at best, impossible at worst. In various ways, we have tried to compromise by allowing that there is knowledge beyond our reach, while recording only acceptable fact. As far as possible, we give references to the original source for all vav grant and unusual records and comment on their acceptability.
Maps
Distribution for each species is shown on one, or more, of the following maps: World, Polar, Aust. and NZ, Aust., NZ, or Tas. We usually present the minimum number of maps in each account: for example, if a species occurs only in NZ, then only that one map is used. However, for many species, distribution (including extralimital range) is shown on a world map and, in more detail on, say, the Aust. and NZ map. All maps have breeding and non-breeding distribution shown in the same manner: breeding distribution is coloured full red. Areas where birds are recorded without known breeding are coloured half-tone red. Vagrant records far from any area of usual occurrence are simply small half-tone red dots or arrows. For islands, and sections of the coast of Antarctica and some coasts elsewhere, breeding and non-breeding are indicated by full red and half-tone red arrows respectively. For seabirds, distribution at sea is in half-tone red without separation into summer and winter or breeding or non-breeding ranges because so little is known of that matter for most species. Where differences between summer and winter range are known it is discussed in the text on distribution or movements rather than shown on a map.
One of the chief difficulties has been to distinguish between breeding and non-breeding ranges, at any rate for species of Aust. and NZ landbirds (see above). When mapping breeding distribution, where ought the lines to be drawn? This can only be a matter of personal judgement, and each person’s judgement will differ. As a general rule, we have tried to outline those areas, based on Atlas blocks, in which the Atlas and other data record breeding as having occurred in the recent past, usually in the past two decades. These areas are coloured full red and include any isolated blocks where breeding was not recorded that are inliers in such areas. The uncertainties of the presentations ought to encourage and give scope to observers to find us at fault.
Movements also were a problem. For species dealt with in this volume, simple migration between breeding and non-breeding areas either does not take place in a clearly defined seasonal manner in our region, except for a few procellariiforms, or movements that do take place are not well enough understood to depict. In short, for the species in Volume 1, it is hardly feasible to present the vagaries of such movement in a succinct form and they are not indicated on maps of distribution.
Much of the information in this section comes from bird reports published in Aust. or NZ. For NZ, annual reports of unusual or interesting records are published as Classified Summarised Notes in Notornis. In Aust., there is no national bird report. However, most States publish or have published bird reports and Corella (formerly Australian Bird Bander) continues to publish accounts of breeding birds in its Seabird Islands Series. These references have generally been abbreviated as follows; each is followed by the year of the report (not the year of publication) except for CSN, which is followed by the volume of Notornis in which it is found.
| Report | Published in or by | |
|---|---|---|
| Qld Bird Rep. | Sunbird (Qld Orn. Soc.) | |
| NSW Bird Rep. | Aust. Birds (NSW Field Orn. Club) (formerly Birds) | |
| Vic. Bird Rep. | Bird Observer’s Club of Australia | |
| Tas. Bird Rep. | Tasmanian Bird Report (Bird Obs. Assoc. Tas.) | |
| SA Bird Rep. | S. Aust. Orn. (SA Orn. Assoc.) | |
| WA Bird Rep. | WA Group of the RAOU | |
| CSN | Notornis (OSNZ) | |
From Vol. 2
The breeding range of a species is the most biologically important part of its total distribution. However, for most species in our region, the limits of breeding and non-breeding distribution are not well known. For many pelagic and colonially nesting species, breeding ranges are often quite well, or even exactly, known but non-breeding ranges are often a total mystery. For terrestrial birds, general occurrence or range may usually be fairly assessed but breeding range and localities are often poorly known.
In compiling texts and maps, a great many sources are used. For species occurring in Aust. and NZ, The Atlas of Australian Birds and Atlas of Bird Distribution in New Zealand formed the basis of most accounts, which were supplemented with information from literature published after the atlases and other, unpublished, sources. Annual bird reports were a valuable source of information on local rarities, expansion of range, annual fluctuations in general abundance, and movements.
The Atlases are fundamentally flawed as regards breeding range and are uncertain and inexact guides because observers were not specifically asked or obliged to search for or record evidence of breeding. Thus, one cannot assess the significance of a breeding record or lack of one in an Atlas block. Is a record of breeding a chance sighting of one nest in five years or records of several nests each year? Similarly, a blank block, or one showing occurrence only, does not necessarily mean that breeding does not take place there, only that it was not recorded because the area was visited only in the non-breeding season, or suitable habitat was not investigated, or for other reasons.
Maps
Presentation of the maps remains as in Volume 1, with breeding areas shown in full red and areas of occurrence where breeding has not been recorded in half-tone red. The maps are gross approximations simply because their scale is so small. The limits of different areas are no more precise on our maps than they are on others, such as those in field guides, though it may be easier to place those limits in relation to towns and well -known localities.
In this Volume we have used a map that shows New Guinea, central and e. Indonesia and some islands of the sw. Pacific Ocean with Aust., or with Aust. and NZ. This obviates the need for a world map for species that occur within the HANZAB region but are also only recorded in parts of New Guinea or Indonesia. However, because we know little of the limits of breeding and non-breeding distribution of species in New Guinea and Indonesia, on these maps distribution has usually been shown in halftone red, giving no indication of breeding range outside the HANZAB region.
From Vol. 3
Vagrant species For species new to Australia and its territories or species listed on the Review List of the RAOU’s Record Appraisal Committee (RAC), non-specimen records must be vetted and accepted by the RAC before a species is included on the Australian list or before a record is considered valid. However, the RAC does not review published records unless they have been submitted to them independently. This creates problems with sight-records published before the establishment of the RAC, which, by and large, have not been
vetted.
We have usually listed as acceptable only those sight- or sound-records that have been accepted by the RAC. However, records of species on the Review List but published before the establishment of the RAC and that include an adequate description of a species are usually listed as acceptable. All early sight-records without description and all sight-records since the establishment of the RAC that have not been submitted to the RAC are listed as unverified or unacceptable.
Many unverified reports of rare or vagrant species are published in the RAOU Newsletter (till Dec. 1990) or Wingspan (in Twitcher’s Corner), or in OSNZ News. These must be considered unacceptable records until they have been submitted to the RAC (Aust.), Rare Birds Committee (RBC; NZ) or relevant State authority. In the accounts these are usually listed as ‘unverified’, without reference. In Aust., records in State bird reports are accepted except for species on the RAC Review List.
Populations
In the Charadriiformes, the estimated total Aust. population for each species is taken from Watkins (1993). We have also included the results of long-term surveys conducted in Aust. and NZ. In Aust., regular counts are usually only a small proportion of the estimated total population. However, we have included these data to show annual variation in numbers in Aust., which, for at least some species, reflects breeding success.
Unlike earlier volumes, Christmas I. refers to the Australian territory in the Indian Ocean unless followed by ‘(Pac.)’, which then indicates the island in the central Pacific Ocean.
Maps
Presentation of maps remains as in Volume 2, with breeding areas shown in full red and areas of occurrence where breeding has not been recorded in half-tone red. Because we know little of the limits of breeding and non-breeding distribution of species in New Guinea and Indonesia, distribution in these regions has usually been shown in half-tone red, giving no indication of breeding range outside the HANZAB region. [this paragraph repeated in Volume 4]
REFERENCES
- Watkins, O. 1993. RAOU Rep. 90.
From Vol. 5
The detailed descriptions of distribution need to be read in conjunction with the maps, which themselves obviate the need for much text. Because the maps can be presented at only a small scale, the text describes the mapped distribution of each species state by state or regionally, indicating whether the species is widespread throughout that range, scattered or patchily distributed; and gaps or continuities in distribution that may not be obvious from the maps are discussed.
The breeding range of a species is the most biologically important part of its total distribution yet for most species in the HANZAB region one of the chief difficulties in assembling both texts and maps is to distinguish between breeding and non-breeding ranges. For most terrestrial species of Aust. and NZ, general occurrence or range may usually be fairly assessed but breeding distribution and localities are often poorly known, even for colonially breeding species. The discussion of breeding distribution relies on the map and the preceding discussion of overall distribution.
In compiling text and maps, a great many sources are used. For species occurring in Aust. and NZ, The Atlas of Australian Birds (Blakers et al. 1984) or The Atlas of Bird Distribution in New Zealand (Bull et al. 1985) form the basis for discussing and presenting both breeding and non-breeding ranges in the text and the maps. These known ranges are supplemented by records published since then (of special note are the Atlas of Victorian Birds [Emison et al. 1987] and Birds of the Australian Capital Territory. An Atlas [Taylor & COG 1992]), some unpublished records and, for Aust., data from the NRS. Annual bird reports for a variety of regions or states are a valuable source of information on local rarities, changes in range, annual fluctuations in general abundance, irruptions, and movements.
However, for Aust., The Atlas of Australian Birds is an imperfect record of breeding distribution because observers were not required to search for, or even submit, evidence of breeding (though it was encouraged); and many areas beyond the well-populated e. coast, SE and SW remained little visited; some areas may never have been visited for more than a few hours and not certainly when breeding may have occurred or in breeding habitat of a species within that block. Thus it is difficult to assess the significance of a breeding record or the lack of one in the Atlas. For all that, however, the Atlas remains the best record of breeding and no n -breeding distribution in Aust. Combined with other sources, the text and maps are thus records of known non-breeding and breeding distribution, though for all but a few species they remain an incomplete record of these ranges. By taking the approach we have we hope to stimulate observation of breeding range and publication of such observations to fill the all-too-obvious gaps in our knowledge. The New Atlas of Australian Birds currently underway
should also provide much useful information.
For NZ, The Atlas of Bird Distribution in New Zealand (Bull et al. 1985) did not distinguish breeding distribution on the maps, stating that ‘apart from pelagic species and migrant waders, most New Zealand species breed throughout their ranges and the exceptions are noted with the relevant maps’. Records of breeding are, however, summarized for each grid square in a microfiche appendix to the Atlas. The NZ Atlas suffers the same problems as the Aust. Atlas regarding recording of breeding distribution. Thus, breeding distribution for NZ is determined from published descriptions and records, and the microfiche records of the NZ Atlas.
Figure 1 shows the regions, divisions and districts of the various Aust. states that are used in conjunction with town and place names and geographical features to describe distribution. The end-paper map for NZ (inside rear cover [see Gazeteer]) shows the regions of the main islands which are used to describe distribution there.
Vagrant and rare species
For species new to Aust. and its territories or species listed on the Review List of the Birds Australia Rarities Committee (BARC, which was formerly the RAOU Records Appraisal Committee [RAC]) , non-specimen records must be vetted and accepted by BARC before a species is included on the Aust. list or before a record is considered valid (see Palliser 1999 and Palliser & Eades 2000 for a copy of the Review List and the role of BARC). However, BARC does not review published records unless they have been submitted to them independently. This creates problems with sight records published before the establishment of BARC or, before that, the RAC, which, by and large, have not been vetted.
For all such species, we have usually listed as acceptable only those sight- or sound-records that have been accepted by BARC. However, records of species on the Review List but published before the establishment of the RAC/BARC and that include an adequate description of a species are usually listed as acceptable. All early sight-records without description and all sight-records since the establishment of the RAC/BARC that have not been submitted to the BARC are listed as unverified or unacceptable.
Many unverified reports of rare or vagrant species are published in the RAOU Newsletter (till Dec. 1990) or Wingspan (in Twitcher’s Corner) , or in OSNZ News. These must be considered unacceptable records until they have been submitted to the BARC or relevant State authority in Aust., or the Rare Birds Committee in NZ. In the accounts these are usually listed as ‘unverified’, without reference. In Aust., records in State bird reports are accepted except for species on the BARC Review List.
Status
The default for status of species and subspecies in Aust. is Garnett (1993), though for a few species we were able to include status from Garnett & Crowley (2000). For status in the various Aust. states, we used Stanger et al. (1998). International status of species came from Collar et al. (1994).
Populations
For passerines of the HANZAB region there are few estimates of total population size, except for many endangered taxa. We have included estimates of densities or relative abundance where they are available, and the results of any long-term surveys conducted in Aust. and NZ.
Maps
Presentation of maps remains as in previous volumes, with breeding areas shown in full red and areas of occurrence where breed ing has not been recorded in half-tone red. Because we know little of the limits of breeding and nonbreeding distribution of species in New Guinea and Indonesia, distribution in these regions has usually been shown in halftone red, giving no indication of breeding range outside the HANZAB region. Maps of distribution appear for all species except:
- extinct species
- those species that receive only brief paragraph treatment in the text (see Treatment and Presentation above)
- failed introductions to the HANZAB region.
We have not tried to show movements on maps.
REFERENCES
- Blakers, M., et al. 1984. The Atlas of Australian Birds. Melbourne Univ. Press, Melbourne.
- Bull, P.C., et al. 1985. The Atlas of Bird Distribution in New Zealand. Orn. Soc. NZ, Wellington.
- Collar, N.J., et al. 1994. Birds to Watch. 2. Birdlife Conserv. Ser. 4. BirdLife Int., Cambridge.
- Emison, W.B., et al. 1987. Atlas of Victorian Birds. Dept. Cons. Forests and Lands & RAOU, Melbourne.
- Garnett, S. 1993. RAOU Rep. 82.
- —, & G. Crowley. 2000. Action Plan for Australian Birds 2000. Environment Aust. , Canberra.
- Palliser, T. 1999. Wingspan 9(3): 27-30.
- —, & D.W. Eades. 2000. Wingspan 10(3): 26-9.
- Stanger, M., et al. 1998. CSIRO List of Australian Vertebrates: A Reference with Conservation Status. CSIRO Publ., Melbourne.
- Taylor, M., & COG (Canberra Orn. Grp) . 1992. Birds of the Australian Capital Territory. An Atlas. Canberra Orn. Grp & National Capital Planning Auth., Canberra.
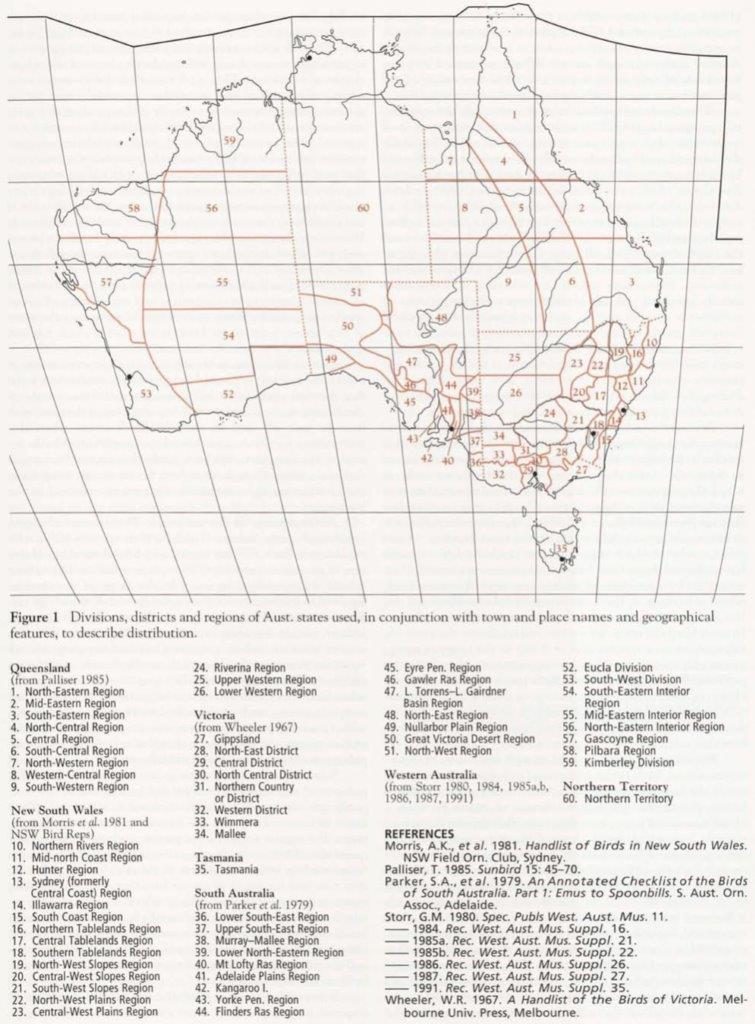
From Vol. 6
Appendix IV [see Fig. 1 from volume 5 above – note extra region, no. 61, added in volume 7 – see revised map in Gazetteer] shows the regions, divisions and districts of the various Aust. states that are used in conjunction with town and place names and geographical features to describe distribution. The end-paper map for NZ (inside rear cover) shows the regions of the main islands which are used to describe distribution there.
From Vol. 7
… A map showing the regions, divisions and districts of the various Aust. states that are used in conjunction with town and place names and geographical features to describe the distribution [see map in Gazetteer] … map for NZ (see map in Gazetteer) shows regions on the main islands which are used to describe distribution there.
CONSERVATION STATUS
This is a new section not in the HANZAB books. There is some overlap with Threats and Human Interactions.
The sources used are referenced using the following abbreviations:
- APAB 2020 refers to The Action Plan for Australian Birds 2020 (Eds ST Garnett and GB Baker) CSIRO Publishing, Melbourne.
- NZTCS 2021 refers to Robertson, Hugh A. , Karen A. Baird, Graeme P. Elliott, Rodney A. Hitchmough, Nikki J. McArthur, Troy Makan, Colin M. Miskelly, Colin. J. O’Donnell, Paul M. Sagar, R. Paul Scofield, Graeme A. Taylor and Pascale Michel (2021) Conservation status of birds in Aotearoa New Zealand,.
New Zealand Threat Classification Series 36. Department of Conservation, Wellington. https://www.doc.govt.nz/about-us/science-publications/conservation-publications/nz-threat-classification-system/ (accessed 15/2/2023)
Where the IUCN status has been updated since these publications, the more current rating has been used.
THREATS AND HUMAN INTERACTIONS
[Note this section included in all volumes online]
From Vol. 5
For this and remaining volumes, we have decided to place all information relating to threats to a species, and the variety of human interactions recorded for a species, in a single place where previously this information was included in the last paragraphs of both Habitat and Distribution and Population. The overall status of species, and subspecies, is still retained in the section on Distribution and Population, immediately preceding this new section. Here we concentrate on summarizing those factors that are known to be or are potentially threatening to a species or subspecies. The information compiled by S. Garnett, G. Crowley and J. Brouwer in their various reports (Brouwer & Garnett 1990; Garnett 1993; Garnett & Crowley 2000) were invaluable in this respect, though few details from Garnett & Crowley (2000) were available at the time of publication of this volume of HANZAB.
The second part of this section considers all aspects of human interactions, ranging from use of modified habitats, such as use of urban areas or farmland and impacts of logging or fires, to more trivial aspects, such as collisions with windows and overhead wires.
Fuller details of the various aspects discussed in this paragraph are often contained within the Habitat or Distribution and Population sections, which need to be read in conjunction with the summaries in this section.
REFERENCES
- Brouwer, J. , & S. Garnett. 1990. RAOU Rep. 68.
- Garnett, S. (Ed.) 1993. RAOU Rep. 82.
- –, & G. Crowley. 2000. Action Plan for Australian Birds 2000. Environment Aust., Canberra.
MOVEMENTS
[Information from volumes 1, 2, 3, 4, 5, and 6,7]
From Vol. 1
This section describes the timing and geography of migration and other long-distance movements rather than orientation or physiology. Each account begins with a brief summary of type of movements, whether migratory (all or most individuals moving between breeding and non-breeding ranges), partially migratory (some individuals migratory, others resident), dispersive (movements apparently random within suitable habitat, though this designation may reflect limited knowledge and mask regular migration by part of the population), resident (most individuals non-migratory though some may move long distances) or sedentary (most individuals not normally moving more than 50 km). Other types of movement include eruptions from breeding areas, irruptions into areas outside the normal range, aberrant migration (movement in the opposite direction to most members of the species), post-fledging dispersal, post-breeding dispersal and moult-migration (movement from a breeding area to a moulting site). The term ‘nomadic’ is generally avoided because it seems to be little more than a cloak for our ignorance, and a vague synonym for ‘dispersive’.
For regular migrants, the direction and timing of departure, movements in the non-breeding season, direction and timing of return to the breeding area and movements in the breeding season are set out separately.
Banding returns are generally summarized separately. We have drawn together all records of movements of more than 100 km for Aust., NZ and Antarctica, except for birds banded by the British Antarctic Survey and those banded by the United States Bird Banding Laboratory within the limits of the book but recovered outside it. For a few species, banding returns have been adequately summarized in the literature; otherwise banding returns are either mapped or summarized thus:
46S51E 03 P U 14 2837
288 CRBPO
(1) (2) (3) (4) (5) (6) (7) (8)
- (1)Coordinates of banding site (degrees latitude, degrees longitude)
- (2) month of banding
- (3) age at banding (J = juvenile, P = pullus, 1 = 1 year-old, 1+ = >1 year old, etc.)
- (4) sex (M = male, F = female, U = unknown)
- (5) number of months between banding and recapture
- (6) minimum distance to location of recovery (great circle)
- (7) direction to location of recovery (degrees from N)
- (8) scheme under whose auspices bird banded (listed below).
Maps
For some species, banding recoveries are presented on maps; the symbols used are listed below. In all cases, recoveries are pooled by grid-cell, the centres of which are shown by the symbol; size of the grid-cell is specified at the foot of each map (e.g. 1°lat. x 1°long.; see below, Fig. 1). Where less than 100 recoveries are mapped, an open circle represents one or more recoveries in a grid-cell. Where more than 100 recoveries are mapped (a percentage [%] symbol appears in the legend), the symbols + and 1 to 9 represent a percentage (%) of the total number of recoveries for that map; i.e. 2 indicates that between 20% and 29% of the total number of recoveries were recorded in that grid-cell.
(2) size of grid cell in degrrees latitude by degrees longitude
(3) when present indicates more than 100 recoveries are mapped and that figures on maps are percentages of the total number of recoveries
(4) banding scheme code (see below)
Fig. 1
Codes for contributing banding schemes are as follows:
| ABBBS | Aust. Bird and Bat Banding Scheme, ANPWS, Aust. | |
| BMRC | Bird Migr. Res. Center, Yamashina Inst. Orn., Japan | |
| BBL | Bird Banding Lab., Fish Wildl. Serv. US, Dept Int., USA | |
| CEMAVE | Centro Estudos Migr. Aves, Brazil | |
| CRBPO | Centre Rech. Biol. Popul. Ois., Natn. Mus. Hist. nat., France | |
| IAC | Inst. Antarct. Chileno, Chile | |
| MAPS | Migr. Animal Pathol. Survey (McClure 1974) | |
| NZNBS | NZ National Banding Scheme, NZDOC, NZ | |
| SABRU | S. Afr. Bird Ringing Unit, Univ. Cape Town, Rep. S. Africa | |
| SOPAN | Stajca Orn. Polska Akad. Nauk, Poland | |
| VH | Vogelwarte Hiddensee, East Germany | |
From Vol. 2
Unfortunately, banding returns were not made available for this Volume and so only published banding data have been presented and there are no maps of banding recoveries. Published long distance banding recoveries are listed in the final paragraph of this section. For the Charadriifonnes, most banding recoveries were available through the Australasian Wader Studies Group and their publication The Stilt. Where appropriate, the results of
radio-tracking studies are also presented in the final paragraph of
this section. Banding summaries, as in Volume 1 [see above] …
From Vol. 3
The text summarizes available information that describes movements in the HANZAB region. For species that move into and out of the HANZAB region, relevant extralimital movements are also described. The patterns of movements of species in this volume vary widely, from species that are sedentary (e.g. Wonga Pigeon Leucosarcia melanoleuca) to those that undertake trans-global migration (e.g. Arctic Terns Sterna paradisaea). Some species display a variety of movement strategies, e.g. Silver Gull Larus novaehollandiae. This volume includes the longest band-recovery known to
date: a South Polar Skua Catharacta maccormicki banded in Antarctica and recovered in Greenland.
Presentation of this section remains much as in Volumes 1 and 2. The first paragraph summarizes the patterns of movements of a species and, where appropriate, the nature of passage. If available, flight-speed is also mentioned. Subsequent paragraphs describe movements in detail, often population by population or, for migratory species, in relation to the breeding cycle. The results of banding, colour-marking and radio-tracking
studies are given in the final paragraphs.
Long-distance banding recoveries (> 100 km) are listed in the final paragraph. These data were provided by the Australian Bird and Bat Banding Schemes (ABBBS) in Canberra and by the Australasian Wader Studies Group (AWSG), including records published in their journal The Stilt. Band recoveries are also listed in Corella and Aust. Bird Bander and,
in some cases, might be more complete than data available from the ABBBS. Where appropriate, summaries of other banding records and the results of radio-tracking studies are also presented.
Banding summaries, as in earlier volumes … [see above]
For some species, banding recoveries are presented on maps. The banding site is shown by a symbol and recoveries of birds from that site shown by a smaller version of the symbol. If there was more than one recovery at a site, the number of recoveries is placed next to the symbol.
From Vol. 4
The patterns of movements shown by species in this Volume vary widely, from species that are sedentary (e.g. Papuan Frogmouth Podargus papuensis) to those that are migratory (e.g. Orange-bellied Parrot Neophema chrysogaster). Few species have been adequately studied using marked birds or at biologically useful scales for understanding species movements. Such studies are needed to deal with major problems with the interpretation of survey, count or presence-absence data; for example, it is often difficult to distinguish seasonal changes in conspicuousness and detectability from movements into and out of an area (this being particularly obvious with some species in this Volume, such as the cuckoos).
As in earlier volumes, a summary of banding recoveries are listed in the final paragraph of this section. Where appropriate, summaries of other banding records and the results of radio-tracking studies are also presented in the final paragraph. However, the format of banding summaries has changed from that used in earlier volumes. In this Volume, banding recoveries are presented in categories of distance from banding site (<10 km, 10-49 km, 50-99 km, ≥ 100 km). For Aust,. these figures are calculated from data supplied by the Australian Bird and Bat Banding Schemes (ABBBS) in Canberra, most of which are summarized in Baker et al. (1997). In some instances, it was necessary to incorporate additional recoveries, typically from the Recovery Roundup section of the journals Corella and Australian Bird Bander (e.g. for Regent Parrot Polytelis anthopeplus). Where a species could have been banded in islands of the sw. Pacific or New Guinea or both, these totals are also incorporated (because they could not easily be extracted from the calculations). Details are provided for all long-distance recoveries (i.e. ≥ 100 km) where they are available. The information on recovery rate in distance categories is excluded from species with recent taxonomic splits and where there is subsequent uncertainty as to the specific identity of banded birds (e.g. Baudin’s Zanda baudinii and Carnaby’s Z. latirostris Black Cockatoos in sw. WA). Recoveries were summed for taxa that have been recently lumped and are presented separately in the ABBBS data (e.g. Australian Ringneck Barnardius zonarius).
For each species, long-distance recoveries are presented in the following summarized form:
Kenmore, se. Qld, to Wewak E., Sepik Province, PNG
(1) (2)
(2825 km, 337°, 3 monthsD, Mar., J, M)
(3) (4) (5) (6) (7) (8)
- (1) Banding site;
- (2) recovery site;
- (3) minimum distance from banding site to recovery site (great circle distance);
- (4) direction from banding to recovery site (great circle);
- (5) number of months elapsed between banding and recovery (D; recovery of dead bird);
- (6) month of banding;
- (7) age at banding, if known (P; pullus; J; juvenile, 1 ; 1 year old, blank; unknown or > 1 year old); and
- (8) sex if known (M ; male, F; female, blank ; unknown).
Where appropriate, recoveries showing site-fidelity are also discussed. Longevity from banding records of wild birds is given (if >12 months). No banding maps are presented in this volume.
REFERENCES
- Baker, B., et al. 1997. Annual Report of Australian Bird and Bat Banding Scheme, 1995-96. Environ. Aust., Canberra.
From Vol. 5
This section describes the timing and geography of migration and other movements, or the lack of them. However, few species have been adequately studied using marked birds or at biologically useful scales for understanding movements by birds in the region. The texts in HANZAB are, for most species, the first attempt to collate and synthesize all information available on movements by bird species using the region, at least as far as landbirds of Aust. are concerned; Keast’s (1968) analysis of movements of Aust. honeyeaters is a notable exception. As with the Habitat section, it has proved difficult to assemble even a general overview of movements let alone a detailed analysis for each species. What is assembled here can
be considered only a first step in a fuller understanding of the patterns of movements of A’asian birds, particularly the landbirds.
What is needed are detailed studies that take the information presented in HANZAB and combine that with analyses of quantitative data from as many sources as possible, especially the data gathered by the thousands of volunteers coordinated by Birds Australia itself, including the data from the Australian Bird Count (ABC), the Nest Record Scheme (NRS), and the
data held in both the original and new Atlases of Aust. birds, as well as the banding and recovery data from the Australian Bird and Bat Banding Schemes (ABBBS). Such a task is well beyond the scope of the HANZAB project. It is, however, pleasing to note that such work is being undertaken by P. Griffioen and his colleagues. Preliminary results (see Clarke et al. 1999) have already provided a much clearer picture of the patterns of movements of some species, and the approach developed by them will greatly advance our understanding of movements of at least some Aust. birds. Detailed studies need also to deal with major problems with the interpretation of survey, count and presence-absence data; for example, it is often difficult to
distinguish seasonal changes in conspicuousness and detectability from movements into and out of all. area.
Beyond the lack of systematic and detailed study of movements, the single biggest difficulty with preparation of this section is the failure of amateur and professional ornithologists to define the terms they use to describe the movements on which they report, and the lack of a standard nomenclature of movements that is widely accepted or applicable to all studies (e.g. see above and Pyke et al. 1989). Thus, the term ‘sedentary’ in one study may correspond with the term ‘resident’ in another, neither of which may correspond with the definitions we have adopted here (see below). However, without a clear statement by all. author of what is meant by the terms used, which is so often in the case in annotated checklists and the
like, there is little we can do other than report the observations. In assembling the texts, we collate and synthesize the available information on a species and in doing so the range of movements of a species may become apparent; we then state the patterns of movements shown by a species as defined by us. However, where no patterns emerge or there is little information, we are often forced to summarize movements as they are
described in the literature without being able to state clearly how that relates to the categories we use.
Nomadism A further problem with definitions of movements of Aust. birds is the widespread and often indiscriminate use of the term ‘nomadic’ to describe movements. Certainly, within the HANZAB region, simple migration between a breeding area and a non-breeding one either appears not to take place in a clearly defined seasonal manner, except for a few species, or the movements that do take place are not well enough understood to explain clearly; the movements of some species do appear to be largely unpredictable. Further, while much movement does, of course, occur, it can be greatly influenced by many factors, not all of which are known or understood, including wet and dry conditions within a species’ range, and local movements (sometimes with seasonal patterns) to take advantage of flowering of plants. The definition or description of much movement observed in Aust. being difficult and our knowledge imperfect, many such movements are called ‘nomadic’. However, the idea that many, or even most, Aust. bird species are ‘nomadic’ has become somewhat of all. overriding paradigm for understanding movements of Aust. birds.
With our work on the Aust. land birds, and the passerines in particular, we are often confronted with claims of nomadism or partial nomadism. This is all too often the situation with annotated lists, checklists, regional reports and the like. Yet, in general, claims of nomadism in many of these and other papers are often based only on apparently unpredictable occurrence at a given locality or anecdotal records, with no understanding, or even in ignorance, of the wider scale patterns of movements that may influence such appearances. Such claims often extrapolate from known occurrence in all. area to making claims about the patterns of movements of the species, which is often not justified on the evidence presented or analysis conducted. Many such claims are often no doubt merely based on previously published claims that a species is nomadic, though this is often not stated. All in all, most examples of use of the terms nomadic and nomadism merely perpetuate a lack of critical thinking of patterns of occurrence and movements of avian species, and, in the words of Stephen Marchant at the start of this series, seem ro be little more than a cloak for our ignorance.
Considerably more study and analysis of the movements of individual species are needed before we can confidently state that they are nomadic. A full discussion of the true nature of claims of nomadism, and the validity of the use of the term, will have to wait till later in the HANZAB series, when the information for all Aust. landbirds is assembled and can be analyzed in its entirety. We have tried as far as possible to avoid the terms nomadic and nomadism. However, we have little choice when trying to summarize the patterns reported in the literature.
Arrangements of the accounts This has not changed significantly since Volume 1, and the texts are reasonably self-explanatory. Each account begins with a brief summary of the type of movements exhibited by a species as defined by us (see below), followed by a summary of the range of movements reported in the literature if they differ from that already given. If appropriate, a brief discussion of seasonal changes in range follows, noting breeding and non-breeding ranges and the seasons in which each is occupied. Details of any geographical variation in nature of movements, timing, routes, etc., if any, are given. The first paragraph then goes on to identify briefly other factors relevant to the overall patterns of movements seen in a species, such as altitudinal movements, associations with flowering plants, and response to droughts or floods, irruptions and the like. Lastly, observations on the nature of passage or other movements are briefly given.
Subsequent paragraphs then go on to describe in detail the patterns of movements observed, and introduced in the first paragraph, discussing geographical variation or seasonal differences in movements, and the factors that might influence them. For regular migrants (or partial migrants), the subsequent sections describe the direction and timing of departure from breeding areas, movements in the non-breeding season, direction and timing of return to breeding areas, and movements in the breeding season. For species that appear largely or wholly resident or sedentary we usually have few problems in summarizing the range of movements. As discussed above, all claims of nomadic movements need to be considered carefully and the true nature of such movements often await verification by more detailed studies.
If there is information available, we also discuss details of dispersal of young, where there are details on movements; details of timing and behaviour of dispersal are usually left to Social Organization and Behaviour (see below).
As stated above, there are few species within the HANZAB region for which a comprehensive description of movements can be prepared and there are no species for which in-depth studies of movements across the range of the species are available. For many species, the paucity of real information on
movements can make it difficult to reach any conclusions on their patterns of movements.
We have defined the following categories of movements:
- MIGRATORY: all or most individuals moving between breeding and non-breeding ranges;
- PARTLY MIGRATORY: some individuals migratory, others resident;
- DISPERSIVE or NOMADIC: movements apparently random within suitable habitat, though this designation may reflect limited knowledge and mask regular migration or other patterns of movements by part or all of the population (see discussion above);
- RESIDENT: most individuals non-migratory though some may move long distances;
- SEDENTARY: most individuals not normally moving more than 50km.
Other types of movement include eruptions from breeding areas, irruptions outside the normal range, aberrant migration (movement in the opposite direction to most members of the species), post-fledging dispersal, post-breeding dispersal and moult migration (movement from a breeding area to a moulting site).
Banding
As in previous volumes, a summary of banding recoveries is given in the final paragraph of this section. … [rest of banding information and references as in Movements, Vol 4 above]
No banding maps are presented in this volume.
From Vol. 6 and 7
[Text similar to Vol 4]
FOOD
[ Information from Vol. 1, 2, 3, 4, 5]
From Vol. 1
The description of food and feeding behaviour is open to many biases and much variation between place, time and individuals. For most purposes, therefore, the generalized description of the diet provided in the first sentence of each food section will be adequate; for no species of wild bird has the acquisition of and requirements for nutrients and energy been completely described. Nevertheless such understanding can only be achieved by numerous studies conducted in as much detail as possible with the techniques available and these are comprehensively summarized in later paragraphs. Until recently, there have been few detailed studies of the diet of A’asian birds. Earlier works gave general descriptions of the gut contents of a few individuals (e.g. Campbell; North; Mathews 1910-27; Lea & Gray 1935, 1935-36; Cleland 1910, 1911; Cleland et al. 1918) but these are of little use in terms of the biology of the species. Only in the last decade have thorough and biologically useful analyses been conducted for individual species or groups of species in our area. More recently, Barker & Vestjens (1989) have listed the foods of Aust. birds using published and their own unpublished data. In the first paragraph feeding behaviour is described, quantitatively where possible. In unquantified studies it should be remembered that unusual feeding behaviour is more likely to be described in the literature. For seabirds and herons, nomenclative descriptions of feeding behaviour have been standardized using the following texts: seabirds: Ashmole & Ashmole (1967), Ainley et al. (1984), Harper et al. (1985); ardeids: Recher et al. (1983). The rest of the text, except the last paragraph, contains detailed descriptions of the diet, including complete lists of food eaten with whatever measures of quantity are available. Such data are undeniably difficult to read but to present it in less detail would severely limit its utility. Methods of describing the importance of different foods, each of which has flaws and merits, include the percentage of the total weight (wet or dry), percentage of the total volume, percentage of the number of items or the frequency of occurrence. Occasionally these measures have been combined into some form of index or presented as raw quantities but, wherever possible, the data have been reanalysed into one of the four measures listed above. Further, size of items is sometimes indicated when available (e.g. length of crustaceans and fish); these are presented in our standard arrangement for simple statistical data: mean (standard deviation; range; sample size) or a subset of this, depending on availability of data. Analyses are prone to numerous forms of bias such as differential rates of digestion, post-mortem breakdown, secondary ingestion (where stomach contents of the prey species are mixed with those of the bird) or incomplete sampling (particularly with regurgitation). These must be borne in mind when interpreting the results. Where possible, precedence has been given to those analyses most likely to reflect the true intake (oesophageal over gizzard samples, multiple-flushed regurgitations over single-flushed regurgitations). Detailed description of diet is largely confined to data collected in Aust., NZ or Antarctica. For several Antarctic and tropical seabirds, however, detailed studies conducted extralimitally (Falkland Is, Aldabra, Hawaii) have been described in full as have studies of trans-equatorial migrants in their non- breeding range. Otherwise, extralimital data have been summarized in less detail.
All scientific names have been checked by authorities at the Museum of Victoria, the Australian Museum, the Western Australian Museum, CSIRO Division of Entomology or the University of Queensland, where possible basing the taxonomic order and nomenclature on the Flora of Australia, the Flora of New Zealand or the Zoological Catalogue of Australia. Vernacular names have been used only for birds and mammals and follow the RAOU List of Recommended English Names or the Zoological Catalogue of Australia, Vol. 5., Mammalia (Canberra: AGPS). The final paragraph includes information on intake, feeding frequency and nutritional physiology.
Abbreviations
A number of abbreviations unique to this section are used in the detailed descriptions of food [those from Vol 2, 3, 4, 5 added and list sorted]:
| % freq. | % frequency |
| % no. | % number |
| % vol. | % volume |
| % wt. | % wet weight |
| A | Autumn |
| ad. | adult |
| ads. | adults |
| excl. | excluding |
| fl. | flowers |
| fru. | fruits |
| imm. | immature(s) |
| incl. | including |
| indet. | indeterminate |
| juv. | juvenile(s) |
| larv. | larva, larvae |
| lvs. | leaves |
| obs. | observations |
| S | Spring |
| Su | Summer |
| sh. | shoots |
| sd., sds | seeds |
| tr. | trace |
| unident. | unidentified |
| W | Winter |
REFERENCES
- Ainley, D.G., et al. 1984. AOU orn. Monogr. 32.
- Ashmole, N.P., & M.J. Ashmole. 1967. Peabody Mus. nat. Hist., Yale Univ. Bull. 24.
- Barker, R., & Vestjens. 1989. The Food of Australian Birds. 1 Cleland, J.B. 1910. Emu 9: 219-26. Cleland, J.B. 1911. Emu 11: 79-95.
- Cleland, J.B., et al. 1918. Dept. agric. NSW Sci. Bull. 15: 1-112.
- Harper, P.C. et al. 1985. BIOMASS Handbook 24.
- Lea, A.H., & J.T. Gray. 1935-36. Emu 35: 63-98, 145-78, 251-80, 335-47.
- Lea, A.H., & J.T. Gray. 1935. Emu 34: 275-92.
- Mathews, G.M. 1910-27. Birds of Australia.
- Recher, H.F., et al. 1983. Colonial Waterbirds Vol 6; 1-10.
From Vol. 2
In the sections for the birds of prey, descriptions of the methods of hunting and attack follow Baker-Gabb (1980).
Hunting methods include:
- Concealed and unconcealed perch-hunting: birds are perched and search an area by sight or sound.
- Low slow-quartering: birds follow a twisting path 1-10 m above vegetation, sometimes remaining stationary in strong winds.
- Transect hunting: similar to low slow-quartering but moving in straight lines, with the height above vegetation varying with species.
- Fast contour-hunting: bird flies usually 1-2 m above the ground or vegetation, generally flushing avian prey that is then caught in the air.
- Soaring and prospecting: bird glides ~20 m above ground, surveying a wide area.
- High-quartering and Hovering: bird quarters an area until prey is sighted, often surveying more thoroughly when hovering begins.
- Flushing prey from cover: bird makes a low pass over prey or suitable habitat, trying to flush prey or expose weaker individuals in flock.
- Ground hunting and stalking: may be done on land or in shallow water; may also occur after an unsuccessful attack.
- Listening: usually in conjunction with other hunting methods, though may be used to detect prey in hollows.
- Mediated flushing and shadowing: raptor follows machinery or other animals, including other hunting raptors, catching flu shed prey.
- Co-operative hunting: two or more raptors of the same species combine to catch prey.
Attack methods:
- Dive attack: raptor drops head-first towards prey, throwing its wings back and feet forward about 1 m. above ground.
- Drop attack: raptor raises wings and descends feet-first, lifting wings still higher and falling faster when near ground.
- Direct flying attack: stationary prey taken in fast flapping flight.
- Tail-chasing: fast flapping flight used to overhaul fleeing prey in the air.
- Stooping: aerial attack where raptor beats its wings a few times, then partly closes wings and speed increases, prey being struck, hit or manoeuvred into a more vulnerable position.
- Glide attack: raptor glides, usually from a perch, on a flat trajectory towards prey.
All scientific names, other than those of birds, were checked against the following references or by authorities at the Museum of Victoria and the National Herbarium of Victoria, or both. Plants: George ( 1981), Cunninghametal. (1 981), Hnatiuk (1990). In vertebrates: Worms: Fauchald (1977), Reynolds & Cook (1976); Molluscs: Vaught et at. (1980 ); Insects: CSIRO (1 99 1). Amphibians and reptiles: Cogger et al. ( 1983). Mammals: Bannister et at. (1988).
Abbreviations
[see Vol 1 above]
REFERENCES
- Baker-Gabb, D.J. 1980. A’asian Raptor Assoc. News 1(4): 9–12.
- Bannister,J.L., et al. 1988. Fauna of Australia. 5. Aust. Govt Publ. Serv., Canberra.
- Cogger, H.G. , et al. 1983. Fauna of Australia. 1. Aust. Govt Publ. Serv., Canberra.
- CSIRO. 1991. Insects of Australia. CSIRO, Melbourne.
- Cunningham, G.M., et al. 1981. Plants of Western New South Wales. NSW Govt Printer, Sydney.
- Fauchald, K. 1977. The Polychaete Worms. Nat. Hist. Mus., Los
Angeles. - George, A.S. (Ed.) 1981-. Flora of Australia. Vols 1-. Aust. Govt Publ. Serv., Canberra.
- Hnatiuk, R.J. 1990. Census of Australian Vascular Plants. Aust. Flora &
Fauna Ser. No. 11, Bureau Flora and Fauna, Aust. Gov. Publ. Serv., Canberra. - Reynolds, J .W., & D.G. Cook. 1976. Nomenclatura Oligachaetologica. Univ. New Brunswick, New Brunswick.
- Vaught, K.C., et al. 1980. A Classification of the Living Mollusca. Am.
Malacologists Inc., Melbourne, Fl.
From Vol. 3
For skuas, jaegers, gulls and terns, descriptions of feeding behaviour (at sea) have been standardized using the terminology of Harper et al. (1985). During preparation and review of the terns (Sterninae), much information was provided by S.J.M. Blaber and attributed to him in the accounts. These data have now been published in Blaber et al. (1995; see below) but it was too late to incorporate the citation in individual reference lists. All scientific names, other than those of birds, were checked against the following references or by authorities at the Museum of Victoria and the National
Herbarium of Victoria, or both. PLANTS: For Aust., Hnatiuk (1990); for NZ, Poole & Adams (1963). GENERAL INVERTEBRATES: Marshall & Williams (1972). CRUSTACEANS: R. Marchant, R. Wilson and G. Poore. MOLLUSCS: Vaught (1989); S. Boyd. INSECTS: CSIRO (1991). ECHINODERMS: T. Stranks. FISH: Paxton et al. (1989), Eschmeyer (1990) and Gomon et al. (1994); T. Bardsley. AMPHIBIANS AND REPTILES: Cogger et al. (1983). MAMMALS: Bannister et al. (1988).
Abbreviations
Some special abbreviations are used in the detailed descriptions of food; these are listed above [Vol 1].
REFERENCES
- Bannister, J.L., et al. 1988. Fauna of Australia. 5. AGPS, Canberra.
- Blaber, S.J.M., et al. 1995. Mar. Ecol. Prog. Ser. 127: 1–13.
- Cogger, H.G., et al. 1983. Fauna of Australia. 1. AGPS, Canberra.
- CSIRO. 1991. The Insects of Australia. CSIRO, Melbourne.
- Eschmeyer, W.N. 1990. Catalogue of the Genera of Recent Fishes. Calif. Acad. Sci., San Francisco.
- Gomon, M.F., et al. (Eds) 1994. The Fishes of Australia’s South Coast. State Printer, Adelaide.
- Harper, P.C., et al. 1985. BIOMASS Handbook 24.
- Hnatiuk, R.J. 1990. Census of Australian Vascular Plants. Aust. Flora
& Fauna Ser. 11; Bureau Flora & Fauna. AGPS, Canberra. - Marshall, A.J., & W.O. Williams. 1972. Textbook of Zoology: Invertebrates. Macmillan Press, Melbourne.
- Paxton, ] .R., et al. 1989. Zoological Catalogue of Australia. 7: Pisces. AGPS, Canberra.
- Poole, A.L., & N.M. Adams. 1963. Trees and Shrubs of New Zealand. R.E. Owen, Gov’t Printers, Wellington.
- Vaught, K.C. 1989. A Classification of the Living Mollusca. Am.
Malacologists Inc., Melbourne, Florida.
From Vol. 4
Remsen & Robinson (1990) present a classification scheme for the foraging behaviour of non-raptorial land birds, discussing search behaviour, attack behaviour, foraging site, food taken and food -handling behaviour. This scheme is particularly useful for descriptions of attack behaviour and we have, as far as possible, standardized our descriptions using the terminology and definitions of Remsen & Robinson (1990; see below). However, it is not possible to categorize simply all forms of search behaviours used by such land birds, and there is considerable overlap between search and attack behaviour (see Remsen & Robinson 1990); search behaviour is more readily categorized for some groups, such as the Falconiformes (see HANZAB 2 and below). Search behaviour is said to end once food or food-hiding substrates have been sighted and attacked. Variables of search behaviour that can be measured include: distance covered per unit time; number of stops per unit time; and number of attacks, including number of attacks per unit time. Birds can move between foraging sites by walking, hopping, jumping, leaping, running, climbing, gliding, fluttering or flying.
ATTACK BEHAVIOUR:
- GLEAN: Pick food items from nearby substrates (including ground) that can be reached without full extension of legs or neck.
- REACH: Completely extend legs or neck upward (Reach-up), outward (Reach-out) or downward (Reach-down) to reach food.
- HANG: Use legs or toes to suspend body below feet to reach food that cannot be reached from any other perched position; includes: Hang-Up, Hang-Down, Hang-Sideways and Hang-Upsidedown.
- LUNGE: Manoeuvres that use rapid movements of legs rather than flight to approach and capture prey beyond range of attack by Reaching.
- PROBE: Insert bill into cracks or holes in firm substrate to capture hidden prey.
- GAPE: Insert bill into substrate as in probe, but open bill to widen opening.
- PULL: Grasp, pull or tear with bill, removing sections of substrate.
- SCRATCH: Dislodge section of substrate with feet; mainly used by ground foraging birds.
- SALLY (includes snatch, hawk, hover-glean, hover, pounce of much literature): Fly from perch to attack a food item on any substrate, eventually returning to same or another perch. Sallying divided into:
- (A) SALLY-STRIKE: Attack in a fluid movement without gliding, hovering or landing, and aimed either at flying prey or stationary substrates.
- (B) SALLYHOVER: Like sally-strike except that bird hovers at the target substrate at end of sally.
- (C) SALLY-POUNCE: Bird lands briefly at end of sally to take food from substrate; food either taken back to perch or eaten on the spot.
- SCREEN: Attack in continuous flight.
- FLUTTER-CHASE: Bird accidentally flushes or dislodges prey from a substrate and then chases prey.
- FLUSHPURSUE: Similar to Flutter-chase except bird uses manoeuvre deliberately to flush prey from hiding places and then pursues flying or falling prey.
For owls (Strigiformes), description of methods of hunting largely follow methods described for raptors in HANZAB 2. For
kingfishers (Alcedinidae, Halcyonidae), description of some searching and fishing behaviour followed Harper et al. (1985),
as described for gulls and terns in HANZAB 3.
We have also used a new heading, Detailed studies, in place of the former headings of Adults or Breeding or Non-breeding. We did this because many detailed studies do not identify the ages of the birds for which data were obtained, and many often combine data from throughout the year. We have under this arrangement identified ages or period of the annual cycle when it is known, usually in the notes describing the data that follow.
All scientific names, other than those of birds, were checked against the following references. Plants: Poole & Adams (1963), Hnatiuk (1990) and, for Eucalytpus and Angophora, Chippendale (1988). Invertebrates: General Invertebrates: Marshall & Williams (1972); Molluscs: Vaught (1989). Insects: CSIRO (1991), Naumann (1993), and, for ants, Taylor et al. (1985) . Fish: Paxton et at. (1989), Eschmeyed1990) and Gommon et al. (1994) . Amphibians and reptiles: Cogger et at. (1983 ). Mammals: Bannister et at. (1988) and West & Menkhorst (1995).
Abbreviations
Some special abbreviations are used in the detailed descriptions of food; these are listed above under Vol 1.
REFERENCES
- Bannister, J.L., et al. 1988. Fauna of Australia. 5. Aust. Gov’t Publ. Service, Canberra.
- Cogger, H.G. , et al. 1983. Zoological Catalogue of Australia. 1: Amphibia
and Reptilia. Aust. Gov’t Pub!. Service, Canberra. - CSIRO. 1991. Insects of Australia. CSIRO Publ. , Melbourne.
- Eschmeyer, W.N. 1990. Catalogue of the Genera of Recent Fishes . Calif. Acad. Sci., San Francisco.
- Gommon, M.F., et al. (Eds) 1994. The Fishes of Australia’s South Coast. State Printer, Adelaide.
- Chippendale, G.M. 1988. Flora of Australia. 19. Eucalyptus, Angophora (Myrtaceae). Aust. Gov’t Publ. Service, Canberra.
- Hnatiuk, R.J. 1990. Census of Australian Vascular Plants. Aust. Flora & Fauna Ser. 11; Bureau Flora & Fauna. Aust. Gov’t Publ. Service, Canberra.
- Marshall, A.J., & W.D. Williams. 1972. Textbook of Zoology: In vertebrates. Macmillan Press, Melbourne.
- Naumann, . 1993. CSIRO Handbook of Australian Insect Names. CSIRO Publ. , Melbourne.
- Paxton, J.R. , et al. 1989. Zoological Catalogue of Australia. 7: Pisces. Aust. Gov’t Publ. Serv., Canberra.
- Poole, A.L., & N.M. Andrews. 1963. Trees and Shrubs of New Zealand. R.E. Owen, Gov’t Printers, Wellington.
- Taylor, R.W., et al. 1985. Fauna of Australia. 2. Aust. Gov’t Publ. Service, Canberra.
- Vaught, K.C. 1989. A Classification of the Living Mollusca. Am. Malacologist 1989.
- West, P. , & P.W. Menkhorst (Eds) 1995. Mammals of Victoria. OUP,
Oxford.
From Vol. 5
This section remains largely unchanged from our original approach. As stated in Volume 1, the description of food and feeding behaviour is open to many biases and much variation between place, time and individuals.
The first paragraph opens with a summary statement of the type of food eaten by the species. This is followed by a summary of feeding Behaviour, analyzed quantitatively where possible and discussing, after a brief introductory summary: size of feeding flocks, feeding associations and feeding territories; feeding heights, sites, and food sources; feeding methods, including search, attack and handling behaviour (see below); differences between sexes and interspecific comparisons; adaptations for feeding; pest status; and drinking behaviour. In unquantified studies it should be remembered that unusual feeding behaviour is more likely to be described in the literature.
Subsequent paragraphs discuss the results of Detailed studies of diet of adult birds, if any, usually with separate paragraphs for individual studies or localities. For each study, we state the location of the study, the sample size, and notes relating to aspects that may affect the analysis or presentation of the data, such as years or seasons in which data were collected, or whether birds were breeding or not.
These paragraphs contain detailed descriptions of the diet including complete lists of food types eaten with whatever measures of quantity are available. Such data is undeniably difficult to read but to present it in less detail would severely limit its utility. Methods of describing the relative importance of types of food, each of which has both flaws and merits, include the percentage of the total weight (wet or dry), percentage of the total volume, percentage of the number of food items or the percentage frequency of occurrence. Occasionally these measures have been combined into some form of index or presented as raw quantities but, wherever possible, the data has been re-analyzed into one of the four measures listed above. Analyses are prone to numerous forms of bias, such as differential
rates of digestion, post-mortem breakdown, secondary ingestion (where stomach contents of the prey species are mixed with those of the bird) or incomplete sampling (particularly with regurgitation). These should be borne in mind when interpreting the results. Where possible precedence has been given to those analyses most likely to reflect the true intake (oesophageal over gizzard samples, multiple-flushed regurgitations over single-flushed regurgitations). It is also worth noting that records of
food of adults during breeding periods may include food for
young that is not consumed by adults themselves.
Following discussion of detailed studies, if any, we then list, with references, anecdotal records of foods eaten. The paragraph is headed either Other records or, where there are no preceding detailed studies of diet, the paragraph starts with the comment ‘No detailed studies’.
Feeding and diet of Young are then discussed separately, though details of roles of parents and any helpers in feeding young or frequency of feeding are usually left to the Social Behaviour or Breeding sections. As with adult data, any detailed studies are discussed first, followed by anecdotal records.
The last paragraph, Intake, discusses dietary physiology, including daily intake of food, and size and nutritional or calorific values of food-items, when information on these aspects of diet have been published.
When summarizing the results of detailed studies or listings of other records, the main breakdown of items, with headings used only as needed, is: Algae, Fungi, Plants, Animals and Other matter (which lists non-food items, such as grit, plastics and so on). Under Plants, three main subsections are used — GYMNOSPERMS, MONOCOTYLEDONS and DICOTYLEDONS; under Animals, the main subsections are the major phyla, except for the Arthropoda, where the major classes are used as the main subsections. Below these subsection headings, families, and genera and species within families, are listed in alphabetical order. Note that for nectar-feeding species, anecdotal records of food eaten often do not specify the actual items eaten when birds are foraging at flowers. Throughout this volume, we have assumed that the food taken is nectar when references report feeding ‘at flowers’ or ‘in flowers’, though obviously the birds may be taking other food-items, such as insects. This problem is not confined to anecdotal observations, but detailed studies usually specify assumptions made. Throughout the series, flowers are not considered a food item unless a references clearly states that the flower was eaten.
In the HANZAB region, several non-nectar sources of carbohydrate are important sources of food for many groups of birds (e.g.
honeyeaters [Meliphagidae]). They include (from Paton 1980):
- HONEYDEW, which is the sugary secretions of nymphal stages of psyllids (Psyllidae), aphids (Aphididae) and coccids (Coccidae); it consists of small polysaccharides with some glucose, fructose or sucrose and almost no protein; it can solidify.
- MANNA, which is the sugary fluid that exudes from damaged plant material (e.g. from insect attacks) and later crystallizes; it consists of c. 60% sugar, 16% water, some ash and 20% pectin and uronic acids.
- LERP, which is the protective covering produced by many Aust. psyllids, and is mostly carbohydrate.
Describing search and attack behaviours Remsen & Robinson (1990) … [see Vol 4 above].
For standard sources used for nomenclature for plants and
animals, see Taxonomy and Nomenclature above.
Abbreviations
Some special abbreviations are used in the detailed descriptions of food; these are listed above [Vol 1].
REFERENCES
- Paton, D.C. 1980. Emu 80: 213-26.
- Remsen, J. Y., & S.K. Robinson. 1990. Studies Avian Biol. 13: 144- 60.
SOCIAL ORGANIZATION AND SOCIAL BEHAVIOUR
[Information from Vol. 1, 2, 3, 4, 5. Vol. 1 and 5 text is similar]
From Vol. 1
These are based on the similar sections of Social Pattern and Behaviour in BWP. The intention is to summarize in two sections observations and studies of each species:
- Social Organization — where information on the main characteristics of society is presented, including types of associations and spatial separation between individuals or groups of birds.
- Social Behaviour — where mutual interactions between individuals are described (see e.g. Tinbergen 1953; Krebs & Dawkins 1984).
The accounts are largely factual but interpretative comments have been included; often they are purely descriptive. Sometimes, very little is known on the species; if so, information has been gleaned from published notes or personal observations of naturalists and ornithologists. When possible, the material has been prepared in consultation with ornithologists having special knowledge of the species concerned. By presenting the facts, the accounts will provide a source for use in future analyses of the behaviour of the species and taxa. When possible, the accounts have been standardized and follow the style given below but some flexibility has been maintained to accommodate details on some species.
SOCIAL ORGANIZATION
This concentrates on the relations between individuals or larger assemblages of birds, and their dispersion within the habitat. For general reviews of this topic, see especially Lack (1954, 1968), Wynne-Edwards (1962), Crook (1965, 1970), McKinney (1973) and Wilson (1975). We begin by giving an impression of the social grouping typical of the species, i.e. gregarious (in family parties, flocks), in pairs or solitary. The rest of the paragraph summarizes seasonal changes from the typical pattern, and gives details of flocks, i.e. size, sex and age composition. The account then proceeds under three subheadings (usually three separate paragraphs): Bonds, Breeding dispersion and Roosting.
BONDS
The type of mating system (the social organization of the breeding unit, especially relating to copulation and breeding cycle) is presented in terms of pair-bonds between individuals and share of parental care taken by birds involved in breeding. Pair-bonds are normally heterosexual; although homosexuality is common in captive birds, it is rare in the wild. Generally pair-bonds have been treated as:
- monogamous
- polygynous
- polyandrous
- promiscuous, when no true pair-bond is established
Within a species these mating systems are not always mutually exclusive.
- Monogamy is the most common system in birds and often involves joint care of the young. The bond between a single male and female can be: seasonal, being for all or part of one breeding cycle, a new partner being obtained next cycle; sustained or long-term, where the bond is kept for a longer period, with or without breaking the association during the non-breeding period; life-long, where the bond is maintained until death or disappearance of partner. Other than in seasonal monogamy, a pair is said to divorce if one member pairs with a new mate when their old mate is known to be alive (Richdale 1951). The general term for pairing with two or more individuals is polygamy.
- In polygyny, during one breeding cycle a male typically bonds with two or more females, each female often caring for her own brood without any help from the male.
- The least common system is polyandry where, during one breeding cycle, a female bonds with two or more males, each male usually caring for his own brood without help from the female. Polygyny and polyandry may be either simultaneous or successive (serial). In polyandrous species, the female often plays the leading part in advertising and courtship and may also be the larger and brighter sex. Some polygamous species form harems where a male and a group of females (mate-defence polygyny) or a female and group of males (mate-defence polyandry) associate together. Others form leks where males (male-dominance polygyny) and occasionally females (female-access polyandry) display and are visited by the opposite sex solely for copulation.
- Promiscuity is when birds pair only for mating. It may be the only type of sexual relation in a species, or it may be an additional feature to one of the mating systems listed above e.g. individuals maintain stable pair-bonds with their mates while having promiscuous matings with other individuals of the opposite sex. Promiscuity may be species-characteristic (frequent and widespread) or casual (only occasionally seen). After discussing pair-bonds, we present information, if available, on sex-ratios, age of first pairing, and stage of the annual cycle when pair-formation starts and ends. A summary is given of contributions of individuals of a pair or family to breeding, which covers the topics of cooperative breeding and parental care. Aust. has a large number of co-operatively breeding species (Ford 1989; Russell 1989) but, as none of the species covered in this volume breed co-operatively, introductory details will be presented in a later volume. Details of division of labour between pairs are sometimes given here; elsewhere it was more appropriate to include these details in the paragraph in Social Behaviour on Relations within family group. Finally any associations outside family group are included e.g. créching.
BREEDING DISPERSION
Though this paragraph mainly focusses on breeding birds, some information is given on non-breeding territories. The species is usually categorized as a solitary or colonial nester, although sometimes it may adopt either. Any details, if known, are given on distances between nests. A species may also be territorial, non-territorial or both (depending on time of year, habitat etc.). A territory may be best defined as ‘any defended area’ (Noble 1939; but see Emlen 1957) occupied exclusively by a single bird, pair or larger social unit. It is usually, but not always, fixed in space, has clearly defined boundaries and ownership is proclaimed with distinctive behavioural displays and vocalizations (Davies 1980). Nearly all information relating to territories, including outside breeding season, is presented at this point. Size, important characteristics of habitat and seasonal changes are detailed, as are activities restricted to territories. For information on functional categories such as feeding territories, breeding territories, and further general details see Lack & Lack (1933), Nice (1941), Hinde (1956), Tinbergen (1957), Davies (1978) and Davies & Houston (1984). If known, data are also given on home-range — the area in which individuals, pairs or groups of birds are active.
ROOSTING
This deals with the roosting (sleeping) and loafing (comfort-behaviour e.g. resting, preening) patterns of the species. Birds may roost or loaf solitarily or communally; nocturnally or diurnally. We then summarize: sites, e.g. protected, unprotected, traditional, temporary; seasonal changes; arrival and departure times from roosts; other factors known to affect patterns, e.g. tidal regime in birds using littoral habitats for feeding. For review of functions of communal roosts see Ydenberg & Priss (1981), Amlaner & Ball (1983).
SOCIAL BEHAVIOUR
This section is largely restricted to describing the interactions between individuals of a species, leaving interpretation to the reader. Causal analyses of motivational factors involved in displays have generally been considered to be beyond the scope of this work. Usually the material has been divided into three main categories (ordinarily presented as three paragraphs) — Agonistic behaviour, Sexual behaviour and Relations within family group. Often little is known and categories have been omitted, combined, or sometimes, when most observations occur at the nest, presented as one paragraph headed Behaviour at site. Generally, much emphasis has been given to describing visual DISPLAYS, these being loosely defined as movements (e.g. those associated with nest-building, comfort-behaviour etc.) that have become specialized (ritualized) signals in social communication (see e.g. Daanje 1950; Tinbergen 1952; Morris 1956; McKinney 1965; Smith 1977; Krebs & Dawkins 1984). Because they are more easily made, observations of displays and behaviour are often based on birds in captivity. This is especially true of the ducks. Some attention is paid to calls and other auditory signals, but generally these are dealt with in the Voice section. Display names follow those given in the major references and are customarily given initial capital letters. Use of the same or similar names for displays in different species does not imply homology. The following outline for the account was adhered to when possible but sometimes it has been inappropriate to present the material in this manner.
The introductory paragraph first gives an idea of how well the species has been studied, and the major references and ornithologists that have contributed to the account. Often references are given only at this point to allow for easier reading by avoiding continual repetition. Thus, the reader can assume that all the material has been extracted from these initial references. This is followed by comments on the ease of observing, and the conspicuousness of, displays; aspects serving to integrate members of a flock e.g. flight-intention signals; lastly, any particular behavioural attributes that are not covered in the three main categories mentioned above e.g. comfort-behaviour.
AGONISTIC BEHAVIOUR
This section concentrates on behaviour associated with conflict, centering on displays and observations related to threat, attack and defence. Birds may compete for food, nesting and roosting sites, and mates. Most aggression occurs within a species but limitations in some resources also lead to aggression between species. The paragraph begins by giving the distance round itself that an individual defends. This may be a small area centred on itself, its mate or family, or it may be a larger territory; it may vary seasonally or according to the activity the bird is performing. This gives the reader an idea of the circumstances in which conflict may arise. Most of the rest of the paragraph describes behaviour related to the passage from threat to fighting, including any appeasement displays to avoid attack, or submissive displays during attack. When threatening, an individual repels or intimidates an opponent without actually fighting. Fighting is used here to denote physical contact between birds. It is rarely used by birds as it carries risks of exhaustion, injury or death, but it is often described as it is conspicuous. Appeasement and submissive displays serve to reduce the aggressiveness of the birds to which the displays are directed. Details are then given on escape behaviour, triumph ceremonies and social dominance. ‘Ceremony’ is the usual term for mutual displays between paired birds, a triumph ceremony being one which follows a successful aggressive encounter. Social dominance behaviour may lead to groupings such as hierarchies and peck-orders, and reduces the likelihood of attack, see e.g. Pulliam & Caraco (1984). Such behaviour when recorded for captive birds should be treated cautiously as it may only arise in conditions of artificial crowding. Then follows a description of any anti-predator behaviour, e.g. distraction displays such as injury-feigning or mobbing. However, sometimes anti-predator behaviour is best placed in the Relations within family group paragraph as it often deals with the communication between parents and young. For more information on aggression see Maynard Smith (1979).
SEXUAL BEHAVIOUR
Signal patterns are most explicit between mates or potential mates during pairing and breeding. This category describes interactions between birds, usually of the opposite sex, during these times. When a number of birds gather to display, it is termed communal. Many of the displays of pair-formation or pair-bond maintenance reduce aggressiveness between partners or potential partners. The term courtship has been used broadly to mean behaviour between the sexes at, or before, the onset of breeding, including that between established pairs or individuals attempting to form pairs. When possible, it is presented more specifically under the headings of Advertising, Pair-formation, Courtship-feeding, Greeting and Allopreening and where and when each of these activities occurs. Advertising displays can have a dual function: that of repelling rivals from territories and attracting a mate to territory. However it is usually treated here rather than in Agonistic behaviour. In pair-formation, where there are complex behavioural features, special headings are sometimes needed: e.g. Water-courtship in the Podicipedidae (grebes), Pursuit-flights in the Anatinae (ducks). Greeting displays are observed when members of a pair meet after temporary separation and are often associated with change-over at the nest. Such displays are often seen in colonially nesting birds. Also mentioned is any special type of searching behaviour performed if a pair loses contact. Sometimes, displays between pairs seem unrelated to courtship and are more extensive than greeting displays, and we have used the heading pair-bond maintenance. In courtship-feeding, generally seen only in pairs, one partner feeds its mate, which usually begs in a manner similar to begging in young. If it occurs in a species, it is most often seen before laying but may continue during incubation (see e.g. Lack 1940; Krebs 1970). The degree to which this activity is ritualized can vary and in extreme cases no food is actually passed between the birds. Allopreening (Cullen 1963) may or may not be ritualized and may be performed by the pair simultaneously, reciprocally or unilaterally. Copulation is usually the final topic covered and includes details on pre- and post-copulatory behaviour, and speed, frequency and conspicuousness of copulation. Any revival of sexual activity outside the breeding season is also given at this point.
RELATIONS WITHIN FAMILY GROUP
This section outlines both behaviour of young and between members of a family, and how this changes over time. Sometimes it includes social aspects between members of a pair that are related to nest-building, incubation and chick-raising and that have not been covered above (in some instances overlaps with material in the Breeding account). The main intention, however, is to describe behaviour associated with feeding of young and aspects of communication between adults and young, and between siblings. Sometimes it has been more appropriate to treat anti-predator behaviour, including the alarm signals given by adults and the responses of young, here rather than in the agonistic paragraph.
REFERENCES
- Amlaner, C.J. Jr., & N.J. Ball. 1983. Behaviour 87: 85-119.
- Crook, J.H. 1965. Symp. zool. Soc. Lond. 14: 181-218.
- Crook, J.H. 1970. Anim. Behav. 18: 197-209.
- Cullen, J.M. 1963. Ibis 105: 121.
- Daanje, A. 1950. Behaviour 34: 48-99.
- Davies, N.B. 1978. Pp. 317-50. In: Krebs & Davies 1978.
- Davies, N.B. 1980. Ardea 68: 63-74.
- Davies, N.B., & A.I. Houston. 1984. Pp. 148-69. In: Krebs & Davies 1984.
- Emlen, J.T. 1957. Ibis 99: 352.
- Farner, D.S. (Ed.) 1973. Breeding Biology of Birds.
- Ford, H.A. 1989. Ecology of Birds.
- Hinde, R.A. 1956. Ibis 98:340–69.
- Krebs J.R., & N.B. Davies (Eds.) 1978. Behavioural Ecology.
- Krebs J.R., & N.B. Davies (Eds.) 1984. Behavioural Ecology.
- Krebs, J.R., & R. Dawkins. 1984. Pp. 380-402. In: Krebs & Davies 1984.
- Krebs, J.R. 1970. Ibis 112: 108-10.
- Lack, D. 1940. Auk 57: 169-78.
- Lack, D. 1954. The Natural Regulation of Animal Numbers.
- Lack, D. 1968. Ecological Adaptations for Breeding in Birds.
- Lack, D., &. L. Lack. 1933. Br. Birds 27: 179-99.
- Maynard Smith, J. 1979. Proc. R. Soc. Lond. B 205: 475-88.
- McKinney, F. 1965. Behaviour 25: 120-220.
- McKinney, F. 1973. Pp 6-21. In: Farner 1973.
- Morris, D. 1956. Behaviour 9:75-113.
- Nice, M.M. 1941. Am. Midland Nat. 26: 441-87.
- Noble, GK. 1939. Auk 56: 263-73.
- Pulliam, H.R., & T. Caraco. 1984. Pp. 122-47. In: Krebs & Davies 1984.
- Richdale, L.E. 1951. Sexual Behaviour in Penguins.
- Russell, E.M. 1989. Emu 89: 61-2.
- Smith, W.J. 1977. The Behaviour of Communicating.
- Tinbergen, N. 1952. Quart. Rev. Biol. 27: 1-32.
- Tinbergen, N. 1953. Social Behaviour in Animals.
- Tinbergen, N. 1957. Bird Study 4: 14-27.
- Wilson, E.O. 1975. Sociobiology, the Modern Synthesis.
- Wynne-Edwards, V.C. 1962. Animal Dispersion in Relation to Social Behaviour.
- Ydenberg, R.C., & H.H.T.L. Priss. 1981. Review of Functions of Roosting. Proc. 5th Ann. Conf. Func. Roosting.
From Vol. 2
SOCIAL ORGANIZATION AND SOCIAL BEHAVIOUR
Arrangement mainly follows that of Volume 1, with the following changes.
- In Volume 1, a summary of the contributions by sexes to the breeding attempt was given towards the end of the paragraph on Bonds. In Volume 2, this is given under a new section heading, Parental care, and is followed by details of dependence of young.
- The introduction to Volume 1 states that nearly all information relating to Territories is presented in the paragraph headed Breeding dispersion. It should be noted , however, that descriptions of the advertising and interactive behaviour related to territorial behaviour are usually found in Agonistic behaviour. The Agonistic behaviour paragraph itself deals mainly with conspecific conflict; details of interspecific conflict are given towards the end of the paragraph, with some material being relevant to Alarm (see below).
- For most accounts in Volume 2, the responses of birds when alarmed by predators or people has been treated differently from Volume 1. Responses not associated with nesting, rearing young, etc. (i.e. mainly outside breeding season) , tend to be found at the end of the paragraph headed Agonistic behaviour, in a section entitled Alarm. Responses associated with nesting and rearing young (i.e. mainly in breeding season) have been placed in the paragraph headed Relations within family group in a section called Parental antipredator strategies. In many records it is unclear whether nests or eggs were present; these observations have usually been placed under Alarm. These sections may also overlap with Flock behaviour in the first paragraph of Social Behaviour but, if the details are important, the material has been cross-referenced. The behaviour of young when alarmed is also included in Relations within family group, in a section called Anti-predator responses of young.
- In the accounts for the rap tors, an additional section, called Aerial Activity, has been added to Social Behaviour. Aerial activity of raptors is a conspicuous feature of their behaviour, and many kinds of aerial displays and behaviours have been recorded. However, the function of these is often unknown or unclear, and may be agonistic and sexual. Most information on aerial activity is discussed in this section but may be cited again in later sections if the known function is relevant to those sections
From Vol. 3
SOCIAL ORGANIZATION AND SOCIAL BEHAVIOUR
Arrangement mainly follows that of Volumes 1 and 2. In the skuas Catharacta, there have been many detailed studies, and additional headings have been used for clarity. Within Aust. pigeons and doves, the function of the Display Flight is unclear and may have both territorial and sexual components. We have placed Display Flights in an additional section, Aerial Activity, in the first paragraph of Social Behaviour (as was done for aerial displays in raptors).
From Vol. 4
SOCIAL ORGANIZATION AND SOCIAL BEHAVIOUR
There have been no major changes between this and previous volumes, though we have made an effort to remove overlap between these sections and the Breeding section, resulting in slight changes in emphasis between the three sections. A few other changes were also made.
In Social Behaviour, COMFORT BEHAVIOUR is now an additional heading at the end of the first paragraph, covering mainly the preening and bathing of groups or individuals, as well as thermoregulatory behaviour when not breeding. Advertising displays, with their possible dual function of advertising and communicating with a mate, and advertising and communicating with rival conspecifics, often appears in the section called Territorial advertising in Agonistic behaviour rather than being placed in Sexual behaviour where it often appeared in earlier volumes. When the function of any particular behaviour or display is largely not known and cannot be classified sensibly as either agonistic or sexual behaviour, then it is usually placed in the first paragraph of the account. Behaviour associated with parents losing interest in, or repelling, their young has often been moved from Relations within family group to Social Organization, where dependence of young and length of time families stay together are discussed at the end of Parental care.
From Vol. 5
SOCIAL ORGANIZATION AND BEHAVIOUR
The aim of the two sections is to summarize observations and studies on:
- the main characteristics of society, including types of associations and spatial separation between individuals or groups of birds (Social Organization)
- mutual interactions between individuals (Social Behaviour).
The accounts are largely factual but interpretative comments have sometimes been included; often the information presented is purely descriptive. When possible, the material has been prepared in consultation with ornithologists having special knowledge of the species concerned. By presenting the facts, the accounts will provide a source for use in future analyses of the behaviour of the species and taxa. When possible, the accounts have been standardized and follow the style given below but some flexibility has been maintained to accommodate details on some species. When very little is known on a species, the two sections may be amalgamated.
SOCIAL ORGANIZATION
This concentrates on the relations between individuals or larger assemblages of birds, and their dispersion within their habitat. For general reviews of this topic, see Lack (1968), Wynne-Edwards (1962) , Crook (1965, 1970 ), McKinney (1973 ), Wilson (1975) and Matthysen (1993). We begin by giving all. impress ion of the social grouping typical of the species, i.e. gregarious (in family parties, flocks), occurring in pairs, or solitary. The rest of the paragraph summarizes seasonal changes from the typical pattern, particularly changes from breeding to non-breeding season, and gives details of flocks, i.e. size and composition (of different sexes and ages ). The account then proceeds under three main subheadings (usually in separate paragraphs): Bonds, Breeding dispersion and Roosting.
Bonds
The type of mating system (the social organization of the breeding unit, especially relating to copulation and breeding cycle) is presented in terms of pair-bonds between individuals, and the share of parental care taken by birds involved in breeding. Unless stated, we are reporting the SOCIAL BOND, which is defined in relation to pairing and not necessarily in relation to the biological parenting of offspring (see below). Pair-bonds are normally heterosexual; although homosexuality is common in captive birds, it is rare in the wild (but see, for example, Silver Gull Larus novaehollandiae, in HANZAB 3). In general, pair-bonds have been classified as:
- monogamous
- polygamous, which its-elf can be further classified as (2a) polygynous or (2b) polyandrous
- promiscuous, when no true pair-bond is established.
Within a species these mating systems are not always mutually exclusive.
- MONOGAMY is the most common system in birds and often involves joint care of the young. The bond between a single male and female can be: SEASONAL, being for all or part of one breeding cycle, a new partner being obtained next cycle; SUSTAINED or LONG-TERM, where the bond is kept for a longer period, with or without breaking the association during the non-breeding period; and LI FE-LONG, where the bond is maintained until the death or disappearance of a partner. Other than in seasonal monogamy, a pair is said to DIVORCE if one member pairs with a new mate when their old mate is known to be alive (Richdale 1951).
- The general term for pairing with two or more individuals is POLYGAMY.
- In POLYGYNY, during one breeding cycle a male typically bonds with two or more females, each female often caring for her own brood without any help from the male.
- The least common system is POLYANDRY where, during one breeding cycle, a female bonds with two or more males, each male usually caring for his own brood without help from the female. Polygyny and polyandry may be either SIMULTANEOUS or SUCCESSIVE (SERIAL). In polyandrous species, the female often plays the leading part in advertising and courtship and may also be the larger and brighter sex. Some polygamous species form HAREMS where a male and a group of females (MATE-DEFENCE POLYGYNY ) or a female and group of males (MATE-DEFENCE POLYANDRY) associate together. Others form LEKS where males (MALE-DOMINANCE POLYGYNY) and occasionally females (FEMALE-ACCESS POLYANDRY ) display and are visited by the opposite sex solely for copulation.
- PROMISCUITY is when birds pair only for mating. It may be the only type of sexual relation in a species, or it may be an additional feature to one of the mating systems listed above, e.g. individuals maintain stable pair-bonds with their mates while having promiscuous matings with other individuals of the opposite sex. Promiscuity may be SPECIES -CHARACTERISTIC (frequent and widespread) or CASUAL (only occasionally seen). In the past, monogamy was accepted as the predominant mating system in birds, appearing to occur in c. 90% of species (Lack 1968 ). Genetic studies have since shown this to be often untrue. Many birds, such as the fairy-wrens Malurus (this volume) have a monogamous social system, where there is social pair-bond between a primary male (often called the breeding male) and a breeding fema le, but have a promiscuous mating system, with many extra-pair copulations taking place (Rowley & Russell 1997 ; see accounts this volume). For reviews on sexual selection, see Emlen & Oring (1977), Andersson (1994) , Ryan (1 997) and Wagner (1998 ). The occurrence of MATE-GUARDING is often given in this paragraph; it is the defence of a female by her mate , most likely to prevent insemination of the female by other males, though other functions have been suggested (see Samson 19 76 ); for reviews, see Mock (1983 ) and Birkhead et al. (1987) . We then present information, if any, on the stage in the annual cycle when pair formation starts and ends. After discussing pair-bonds, we discuss SEX-RATIOS in population and any differences with age or over time; and AGE OF FIRST PAIRING and of FIRST BREEDING. Then follows a discussion of Co-operative breeding in those species that breed co-operatively (which may include discussions of the previous two points). Co-operative breeding species are characterized by the involvement of individuals other than the breeding pair. Typically, young adults that are old enough to disperse and breed independently delay dispersal and stay with their natal group as helpers or auxiliaries, for months or years, assisting the breeding pair in the care of young and territorial defence (Skutch 1961; Rowley 1965, 1981; Brown 1978; Woolfenden & Fitzpatrick 1984 ; Ro wley& Russell 1997).
- A pair and their helpers usually form a social unit that stay together year-round on a territory (Rowley & Russell 1997). Often in co-operatively breeding groups only one female lays (SINGULAR BREEDING) but sometimes two females in a group maintain separate nests (PLURAL BREEDING ) (Brown 1987). (While the term communal breed ing is sometimes considered synonymous with co-operative breeding, it is sometimes restricted to groups in which two or more females lay in the same territory [plural breeding], whether in one or several nests, and assist each other to rear their broods [Skutch 1987]). Most arguments regarding the occurrence of co-operative breeding focus on the idea that the helpers are in some way related to the offspring they help rear, or that they gain some other benefit for themselves, such as breeding experience, access to mates or territories, or improved survival through protection from predators.
To analyze how helping behaviour is affected by kinship, DNA fingerprinting is now often used. For some discussions on the possible benefits of co-operative breeding, see Woolfenden & Fitzpatrick (1984) , Jamieson & Craig (1987), Emlen (1991), Clarke (1 995), Dunn et al. (1995) and Rowley & Russell (1997). There are a large number of co-operatively breeding species in Aust., with co-operative breeding recorded in at least 85 species, of which 74 are passerines (Rowley & Russell 1997). There are various ideas as to why co-operative breeding is so common in Aust. (e.g. Rowley 1968, 1976; Dow 1980; Ford et al. 1988; Russell 1989; Russell & Rowley 1993 ; Cockburn 1996; Rowley & Russell 1997). In the species accounts, a summary is given of data on co-operative breeding, including an outline of contributions by individuals of the breeding group to the breeding attempt. The details of rates of contributions by individuals are usually given in Breeding or Food. Towards the end of this paragraph we usually summarize the roles of parents in the breeding attempt, sometimes under the heading Parental care if there is much information that relates to social organization. However, in an attempt to reduce overlap between sections, often all details are now only presented in the Breed ing section, with additional information to be found in Social Behaviour (Relations within family group) .
For the same reason, information on the timing of independence of young, which often indicates the breakdown in bonds between parents and offspring, is also given in Breeding. Finally, this paragraph discusses dispersal of young from, and philopatry to, natal areas, as they are often implicated in discussions on social organization, particularly co-operative breeding (e.g. Emlen 1982, 1991; Greenwood & Harvey 1982 ; Brown 1987; Pruett-Jones & Lewis 1990; Stacey & Ligon 1991; Koenig et al. 1992); dependence of young and length of times families stay together are discussed, though descriptions of behaviours between adults and young, including behaviour associated with parents losing interest in or repelling their young are usually given under Relations within family groups (in Social Behaviour). The discussion of dispersal of young here often needs to be read in conjunction with the Movements section and, occasionally, it is more appropriate to place the whole discuss ion in Breeding dispersion (in Social Organization) or in the Movements section. Associations outside family group, such as creching, are also given in this paragraph.
Breeding dispersion
This paragraph mainly focuses on distances between active nests or breeding birds, but information on all types of territories and dispersion is included. Species are usually classed as solitary or colonial nesters, though sometimes a species may adopt either strategy. Any details, if known, are given on distances between nests. A species may also be territorial, non-territorial or both (depending on time of year, habitat, etc.) . A TERRITORY is often defined as any defended area (Noble 1939; Hinde 1956; Brown & Orians 1970; but see Emlen 1957), occupied exclusively by a single bird, pair or larger social unit. It is usually, but not always, fixed in space, has clearly defined boundaries, and ownership is proclaimed with distinctive behavioural displays and vocalizations (Davies 1980). Different types of territories have been classified (see Hinde 1956). Details of territories are given in this paragraph, including size, important characteristics of habitat, seasonal changes and activities restricted to territories. The term territories is often used loosely or uncritically by researchers (for examples, see Pyke et al. 1996); others use alternative terms, such as exclusive home-ranges, that imply that it is not known if the area is defended or not. Where practical, we use whatever term a researcher has applied. Many studies of territories relate to breeding territories, but some groups, such as the honeyeaters (Meliphagidae; see Pyke et at. [1996]), often form FEEDING TERRITORIES and these are also discussed here. In many species, feeding territories and HIERARCHIES are closely related and both are important to the discuss ion on social organization . Thus, details of hierarchies are often given here, though for some species (and in previous volumes ) they were discussed in Social Behaviour under the heading Social dominance. Hierarchies noted for captive birds should be treated cautiously as they may only arise in conditions of artificial crowding. Territories and hierarchies often relate to birds within a species, but limitations in some resources can mean that other species are involved in forming INTERSPECIFIC DOMINANCE HIERARCHIES or defend species from other species. Finally, if known, data are given on home-range, which is the area in which individuals, pairs or groups of birds are active. For information on functional categories of territories, such as feeding territories, breeding territories, and for some further background, see Lack & Lack (1933 ), Nice (1941), Hinde (1 956) , Tinbergen (1957) , Dav ies (1978) , Davies & Houston (1984), Carpenter (1987) and Pyke et al. (1996) . Size of territories may be influenced by population density (Krebs 1971; Myers et al. 19 79) , abundance of food
(Franzblau & Collins 1980) and habitat structure (Smith & Shugart 1987). It should be noted that descriptions of the actual behaviours related to advertising and defending territories are usually found in Social Behaviour.
Roosting
This deals with the pattern of roosting (sleeping) and loafing (comfort behaviour, e.g. resting, preening) of a species. Birds may roost or loaf solitarily or communally; and nocturnally or diurnally. We then summarize: sites (e.g. protected, unprotected, traditional, temporary), seasonal changes, times of arrival and departure from roosts, and other factors known to affect patterns (e.g. tidal regime in birds using littoral habitats for feeding). For reviews of functions of communal roosts, see Amlaner & Ball (1 983 ), Weatherhead (1983 , 1987) Ydenberg & Prins (1984) and Zahavi (1 996) .
SOCIAL BEHAVIOUR
This section is largely confined to describing the actual behaviours that occur in interactions between individuals of a species. Causal analyses of motivational factors involved in displays have generally been considered to be beyond the scope of this work. For some general references on social signals and behaviours in birds, see Tinbergen (1964) , Krebs & Dawkins (1984) , Brown (1 994), Johnstone (1997), and Duttmann et al. (1998). Usually the material has been divided into three main categories (ordinarily presented as three paragraphs): Agonistic behaviour, Sexual behaviour and Relations within family group. Behaviours are usually only designated as agonistic or sexual if the sources of the information have stated thus. When the function of any behaviour or display is not known and cannot be classified sensibly as either agonistic or sexual behaviour, then it is usually placed in the first paragraph of the account. In some behaviours where there is overlapping function, then the behaviour is usually placed in the category to which it seems most relevant, and a cross-reference made in the other category. Advertising displays are examples of behaviour that can be agonistic (communicating with rivals), sexual (communicating with a mate) or both, and sometimes cannot validly be attributed to either. In general, much emphasis has been given to describing visual displays, these being loosely defined as movements that have become specialized (ritualized) signals in social communication (see, for example, Daanje 1950; Tinbergen 1952; Morris 1956; McKinney 1965 ; Smith 1978; Krebs & Dawkins 1984; Johnstone 1997). Because they are more easily made, many observations of displays and behaviour are made on birds in captivity. In the accounts we have tried to keep observations made in the wild and those made in captivity separate. Some attention is paid to calls and other auditory signals, particularly in the passerine volumes, but usually these are dealt with in the Voice section (the capitalized names of vocalizations in Social Behaviour indicate that some description of the sound and its use will be found in Voice). Names of displays follow those given in the major references and are customarily given initial capital letters. Use of the same or similar names for displays in different species does not imply homology. Equally, studies of related species by different researchers have often used different names for what appear to be identical displays with identical functions. The following outline for the account was adhered to when possible but sometimes it has been inappropriate to present the material in this manner.
The introductory paragraph first gives an idea of how well the species has been studied, and lists the major studies, and where they were conducted, on which the account is based. Often references are given only at this point to allow for easier reading by avoiding continual repetition. Thus, the reader can assume that all the material has been extracted from these initial references, or that material from a particular locality has been extracted from the identified source. This is followed by comments on the ease of observing, and the conspicuousness of, displays. Sometimes the heading Flock behaviour is used to describe behaviours that serve to integrate members of flocks, e.g. flight -intention signals or use of contact calls by feeding flocks. We then give any particular behavioural attributes that are not covered in the three main categories mentioned above. Lastly, comfort behaviour is summarized, covering mainly preening and bathing of groups or individuals, as well as thermoregulatory behaviour when not breeding.
Agonistic behaviour
This section concentrates on behaviour associated with conflict, centring on displays and observations related to threat, attack and defence over food, nesting and roosting sites, and mates. This paragraph deals mainly with aggress ion within a spec ies. It firstly describes behaviour related to TERRITORIAL ADVERTISING (communicating with rivals), then is followed by the escalation of aggression from threat to fighting, including any appeasement displays to avo id attack or submiss ive displays during attack. When threatening, an individual repels or intimidates an opponent without actually fighting. Fighting is used here to denote physical contact between birds; it is rarely used by birds as it carries risks of exhaustion, injury or death, but it is often described as it is conspicuous. Appeasement and submissive displays serve to reduce the aggressiveness of the birds to which the displays are directed. Details are then given on escape behaviour and triumph ceremonies. Ceremony is the usual term for mutual displays between paired birds, a triumph ceremony being one which follows a successful aggressive encounter. In the passerines, Social dominance is often dealt with in territories in Social Organization, though descriptions of social signals remain here. Usually, after dealing with conspecific conflict, any interesting
descriptions of behaviours given during interspecific conflict are noted. The paragraph finishes with Alarm where descriptions are given of responses to predators, such as mobbing, which are not associated with nesting or rearing young (i.e. mainly outside breeding season) . Responses during a breeding attempt are covered in the paragraph Relations within
family group. For many records it is not clear whether nests or eggs are present, and these observations are usually placed under Alarm. For more background information on aggression, see Maynard Smith (1979).
Sexual behaviour
Signal patterns are most explicit between mates or potential mates during pairing and breeding. This category describes interactions between birds, usually of the opposite sex, during these times. When a number of birds gather to display, it is termed communal. Many of the displays of pair-formation or pair-bond maintenance reduce aggressiveness between partners or potential partners. The term courtship has been used broadly to mean behaviour between the sexes at, or before, the onset of breeding, including that between established pairs or individua ls attempting to form pairs. When possible, it is presented more specifically under the headings of Advertising, Pair-formation, Courtship feeding, Greeting and Allopreening, discussing where and when each of these activities occurs. Advertising displays are placed here if it is thought they are more sexual in function than they are advertising by rival birds. In Pair-formation, where there are complex behavioural features, special headings are sometimes needed. Greeting displays are observed when members of a pair meet after temporary separation and are often associated with change-over at the nest. Such displays are often seen in colonially nesting birds. Also mentioned is any special type of searching behaviour performed if a pair loses contact. Sometimes, displays between pairs seem unrelated to courtship and are more extensive than greeting displays, and we have used the heading Pair-bond maintenance. Courtship feeding is taken to be the feeding of one member of an adult pair by the other. It is often associated with copulation and laying but sometimes continues after this, and in some species seems to have no relationship to copulation at all (Campbell & Lack 1985 ; also see, for example, Lack 1940, and Krebs 1970). The degree to which this activity is ritualized can vary and in extreme cases no food is actually passed between birds. Allopreening (Cullen 1963) mayor may not be ritualized and may be performed by the pair simultaneously, reciprocally or unilaterally. Copulation is usually the final topic covered and includes details on behaviour before and after copulation, and speed, frequency and conspicuousness of copulation. Any revival of sexual activity
outside the breeding season is also given at this point. The term mating is often found in the literature; often it is clear that the author means copulation but at other times exactly what has taken place is not known and such observations are usually omitted.
Relations within family group
This section outlines the social behaviour of young within the family, the social development of young and interactions between young and their parents. Sometimes it includes social aspects between members of a pair that are related to feeding and caring for nestlings and fledgelings that have not been covered above (in some instances overlaps with material in the Breeding section). The main intention, however, is to describe behaviour associated with feeding of young and aspects of communication between adults and young, and between siblings. Alarm responses of young have been given under the heading Anti-predator responses of young. Alarm responses of adults associated with nesting and rearing young have been placed under the heading Parental anti-predator strategies.
REFERENCES
- Amlaner, C.J. Jr, & N.J. Ball. 1983. Behaviour 87: 85-119.
- Andersson, M. 1994. Sexual Selection. Princeton Univ. Press, Princeton,
New Jersey. - Bateson, P. , & P. Klopfer. (Eds) 1987. Perspectives in Ethology. 7. Plenum Press, New York.
- Birkhead, T.R., et al. 1987. Behaviour 101: 101-38.
- Brown, J.L. 1978. Ann. Rev . Ecol. System. 9: 123-55.
- — 1987. Helping and Communal Breeding in Birds. Princeton Univ.
Press , Princeton, New Jersey. - — 1994. Condor 96: 232-43.
- —, & G.H. Orians. 1970. Ann. Rev. Ecol. Syst. 1: 239-62.
- Brush, A.H., & G.A. Clark. (Eds) 1983. Perspectives in Ornithology.
Cambridge Univ. Press , Cambridge. - Campbell , B., & E. Lack. (Eds) 1985. A Dictionary of Birds. T. & A.D. Poyser, Calton, England.
- Carpenter, F.L. 1987. Am. Zool. 27: 223-8.
- Clarke, M.F. 1995. Corella 19: 73-90.
- Cockburn, A. 1996. Pp 21-42 In: Floyd et al. 1996.
- Crook, J.H . 1965. Symp . Zool. Soc. Lond. 14: 181-218.
- — 1970. Anim. Behav. 18: 197-209.
- Cullen, J.M . 1963 . Ibis 105: 121.
- Daanje, A. 1950. Behaviour 34: 48-99.
- Davies, N.B. 1978. Pp 317-50 In: Krebs & Davies 1978.
- — 1980. Ardea 68: 63-74.
- —, & A. I, Houston. 1984. Pp 148-69 In: Krebs & Davies 1984.
- Dow, D.O. 1980. Emu 80: 121-40.
- Dunn, P. O., et al. 1995. Proc. R. Soc. Lond. (B) 259: 339-43.
- Duttmann, H., et al. 1998. Oxford Orn. Ser. 8: 223-46.
- Emlen, J.T. 1957. Ibis 99: 352.
- Emlen, S.T. 1982. Am. Nat. 119: 29-39.
- — 199 1. Pp 301-37 In: Krebs & Davies 1991.
- —, & L.W. Oring. 1977. Science 197: 215- 23.
- Evans, P.R ., et al. (Eds) 1984. Coastal Waders and Wildfowl in Winter.
Cambridge University Press, Cambridge. - Farner, D.s. (Ed.) 1973. Breeding Biology of Birds. National Acad. Sci., Washington, D.C.
- Floyd, R. , et al. (Eds) 1996. Frontiers in Population Ecology. CSIRO Publ. , Melbourne.
- Ford , H.A., et al. 1988. Behav. Ecol. Sociobiol. 22: 239-49.
- Franzblau, M.A., & J. P. Collins. 1980. Oecologia 46: 164-70.
- Greenwood, P.J., & P.H. Harvey. 1982. Ann. Rev. Ecol. Syst. 13: 1-21.
- Hinde, R.A. 1956. Ibis 98: 340-68.
- Jamieson, I.G ., & J.L. Craig. Pp 79-98 In: Bateson & Klopfer 1987.
- Johnstone, R.A. 1997. Pp 155-78 In: Krebs & Davies 1997.
- Koenig, W.O., et al. 1992. Q. Rev . Biol. 67: 111- 50.
- Krebs, J.R. 1970. Ibis 112: 108-10.
- — 1971. Ecology 52: 2-22.
- —, & N.B. Davies. (Eds) 1978. Behavioural Ecology. First edn. Blackwell, Oxford.
- —, — (Eds) 1984. Behavioural Ecology. Second edn. Blackwell, Oxford.
- —, — (Eds) 1991. Behavioural Ecology. Third edn. Blackwell, Oxford.
- —, — (Eds) 1997. Behavioural Ecology. Fourth edn. Blackwell, Oxford.
- —, & R. Dawkins. 1984. Pp 380-402 In: Krebs & Davies 1984.
- Lack, D. 1940. Auk 57: 169-78.
- — 1968. Ecological Adaptations for Breeding in Birds. Methuen, London.
- —, & L. Lack. 1933. Br. Birds 27: 179- 99.
- Matthysen, E. 1993. Current Orn. 11: 93-141.
- Maynard-Smith, J. 1979. Proc. R. Soc. Lond . (B) 205: 475-88.
- McKinney, F. 1965. Behaviour 25 : 120-220.
- — 1973. Pp 6-21 In: Farner 1973.
- Mock, D.W. 1983. Pp 55-84 In: Brush & Clark 1983.
- Morris, D. 1956. Behaviour 9: 75- 113.
- Myers, J.P. , et al. 1979. Auk 96: 55 1-61.
- Nice, M.M.1941. Am. Midland Nat. 26: 44 1-87.
- Noble, G.K. 1939. Auk 56: 263-73.
- Pruett-Jones, S.G ., & M.J. Lewis. 1990. Nature 348: 541-2.
- Pyke, G. H. , et al. 1996. Aust. J. Zool. 44: 297-317.
- Richdale, L.E. 1951. Sexual Behaviour in Penguins. Univ. Kansas Press, Lawrence, Kansas.
- Rowley, I. 1965. Emu 64: 251-97.
- — 1968. Bonn. Zool. Beitr. 19: 362-68.
- — 1976. Int. Om. Congr. Proc. 16: 657-66.
- — 1981. Z. Tierpsychol. 55 : 228-67.
- —, & E. Russell. 1997. Fairy-wrens & Grasswrens. OUP, Oxford.
- Russell, E.M. 1989. Emu 89: 61 -2.
- —, & I. Rowley. 1993. Aust. J. Zoo I. 41: 475-505.
- Ryan, M.J. 1997. Pp 179-202 In: Krebs & Davies 1997.
- Samson, F.B. 1976. Auk 93: 477-97.
- Sebeok, T.A. (Ed.) 1977. How Animals Communicate. Indiana Univ. Press, Bloomington.
- Skutch, A.F. 1961. Condor 63: 198-226.
- — 1987. Helpers at Birds’ Nests. Univ. of Iowa Press , Iowa City.
- Smith, S.M. 1978. Am. Nat. 11 2: 571-82.
- Smith, T.M., & H.H. Shugart. 1987. Ecology 68: 695-704.
- Smith, W.J. 1977. Pp 545-74 In: Sebeok 1977.
- Stacey, P.B., & J.D. Ligon. 1991. Am. Nat. 137: 83 1-46.
- Tinbergen, N. 1952. Q. Rev. BioI. 27: 1-32.
- — 1957. Bird Study 4: 14-27.
- —1964. Social Behaviour in Animals. Second edn. Chapman & Hall, London.
- Wagner, R.H. 1998. Orn. Monographs 49: 123- 45.
- Weatherhead, P.J . 1983. Am. Nat. 121: 237-43.
- — 1987. Anim . Behav. 35: 614-15.
- Wilson, E.O. 1975 . Sociobiology. Harvard Univ. Press, Cambridge, Massachusetts.
- Woolfenden, G.E. , & J.W. Fitzpatrick. 1984. The Florida Scrub Jay. Princeton Univ. Press, Princeton, New Jersey.
- Wynne-Edwards, V.c. 1962. Animal Dispersion in Relation to Social
Behaviour. Oliver & Boyd, Edinburgh. - Ydenberg, R.C., & H.H.T. Prins. 1984. Why do birds roost communally?
In: Evans et al. 1984. - Zahavi, A. 1996. Behav. Eco!. 7: 118-19.
VOICE
[Information from Vol. 1, 2, 3, 4, 5, 6, 7]
From Vol. 1
A full introduction to this section is deferred until a later volume because at that stage a text will be needed to do justice to the detailed problems of dealing with descriptions of voice for true songbirds. Then we intend to deal with developments in methods and equipment for sound-recording and the proper documentation and storage of reference material; we shall give much closer treatment to details of analysis and quantification of sounds and hope to review, briefly, matters concerning production and reception of sound in the bird and the rapidly advancing knowledge of the importance of sound in the behaviour of birds. For the present it is recommended that the reader consult appropriate sections in Bennet-Clark (1981), Campbell & Lack (1985) or the references listed below, for clarification on many matters and, for more technical details, the excellent account given in the revised Introduction to the section on ‘Voice’ in Volume 5 of BWP (pp 21-32). We have prepared much of this present section from these sources.
The vocalizations of birds have been of interest for a long time. The history of human interest in bird songs and calls dates back to earliest literature and is discussed in detail by Armstrong (1969). However, the scientific study of vocalizations of birds has developed only recently. Recent advances are almost solely due to the development of light-weight recording equipment (particularly tape-recorders [reel-to-reel and cassette] and directional microphones) and the development of the audio spectrograph, which allows us to print a visual impression of a sound (see later). All the same, there has been little scientific study of the vocal behaviour of birds or of the role of sounds in their life-histories in A’asia and Antarctica; there is much work to be done. It is lamentable that few observers take any serious interest in bird calls, though today good quality sound-recording equipment is readily available. We know little about most of the species covered in Volume 1 but many are better known than species to be covered in future volumes. As said in BWP (Vol. 5), ‘Comprehensive and critical studies exist as yet only for a small minority. Most authors have felt compelled by this situation to confine themselves to piecemeal assembly of odd scraps of information, too often merely copied, with variations, in one work after another.’ We have experienced the same difficulties but hope that by presenting in many cases what meagre data we have been able to assemble it will be much easier to see where critical observation should be profitable in future.
Primarily, vocalizations of birds are controlled by the syrinx, which is located at the base of the neck and in the thoracic cavity at the junction where the trachea divides to enter the two bronchi that in turn lead to the lungs. The syrinx varies in complexity but is generally bony and cartilaginous with membranes that are set in vibration by the passage of air. Some groups of birds have specialized modifications associated with the syrinx in the form of swellings on the trachea. Notable among birds in this Volume is the presence of a tracheal bulla, often very complex, which is important in production of sounds in males of many Anseriformes. The mechanisms of the syrinx and sound production are still imperfectly known and much work remains to be done (see Greenewalt 1968; Warner 1972; Brackenbury 1982; Gaunt 1983; Gaunt & Gaunt 1985; Nowicki 1987; Fletcher 1988).
The functions of vocal signals are an integral part of the social organization and behaviour of many species and we are beginning to realize the importance of vocal signals and communication. In field identification, the usefulness of sounds is often underrated but has become increasingly important, e.g. to ecologists engaged in surveys. In nocturnal species, vocalizations are often well developed and have an important role, often replacing visual displays (e.g. in many Procellariiformes, Strigiformes). They are used for communication within and between species and can have different meanings in different circumstances. However, functional analysis of vocalizations is complicated and has been attempted for only a few species in our region. It is often difficult to ascribe a function to particular sounds and we have avoided doing so unless there is good evidence. It can be said, however, that vocalizations function to:
- attract mates
- repel conspecifics, especially in the circumstances of territorial behaviour and defence
- allow individual members of pairs or parents and chicks to recognize one another
- maintain pair-bonds
- attune partners or a group to one another.
Vocalizations have been shown to be important in some very specific circumstances, e.g. playback of male vocalizations stimulated laying in captive Budgerigars Melopsittacus undulatus (Brockway 1962, 1965).
VOCALIZATIONS, SONGS AND CALLS
Vocalization is a term that covers all vocal sounds produced by a bird, both calls and songs. The difference between calls and songs is somewhat artificial and arbitrary. We use these terms rather loosely, ‘call’ generally for the more simple and stereotyped vocalizations of most species; however, we use ‘song’ for a number of species (or groups) where we felt it more appropriate. Calls have been defined as usually short, fairly simple vocalizations, usually used by both sexes, throughout the year and often given in response to particular stimuli. Songs are generally more complex and longer vocalizations and are generally restricted to males during the breeding season, though are sometimes given by females; delivery is periodic, can occur spontaneously and is often given from particular perches or sites and associated with specific postures (Catchpole 1982; Campbell & Lack 1985; Konishi 1985). The term ‘song’ is often used only for song of passerines (Konishi 1985). In this sense, song is a relatively complex pattern of utterances that may be repeated exactly, and so is recognizable not only at the level of species but often at those of groups and individual (BWP). Individuals have a repertoire of distinctive songs or calls that may range from one song or call to many hundreds. Songs or calls are distinct combinations of phrases, notes or syllables; phrases are made up of syllables that are discreet units of a song or call that may be repeated within or between songs; syllables may be individual notes or a combination of notes. Notes (sometimes called elements) are defined as a continuous trace on a sonagram (see later) and are the main units of songs. Notes vary enormously in structure and frequency (pitch; see later). A song-bout is a period of continued singing.
DESCRIPTIONS OF SOUNDS
Sounds of birds are difficult to put into words. There are several components to the vocalizations of birds that need to be described. Frequency (pitch); amplitude (loudness); rhythm and tone or timbre (the quality of sounds). The latter is difficult to convey but requires words to indicate purity, harshness, sweetness, nasal quality; ‘reedy’ or ‘flute-like’ and so on. These can be important descriptions because tonality is more difficult to interpret from sonagrams (discussed later) than the other characteristics, except with experience, but it can be crucial to a good description of voice. Tonal quality is the means by which we can recognize a violin from a piano, an oboe from a clarinet. It is complex and concerns, among several factors, the nature of the harmonic structure of the sounds and often the transients or brief initiating noises that precede and may follow the steady state of a particular sound. Words simply cannot do justice to all of these matters and that is why the objective understanding of bird vocalizations had not made much progress until it was possible to record their utterances, so that they could be played back again and again, and until the sounds could be quantified in some way. The application of frequency spectrum analysis has been the means by which insight has been gained.
Phonetic renderings of sounds are a crude and subjective attempt to convey the quality and sound of a vocalization; they are often quite inadequate in dealing with complex songs, and even more so where there is much variation. Nevertheless, any competent descriptions accompanied by syllabic renderings can tell us something useful about the pattern, pitch, and tone of a vocalization. Onomatopoeic renditions are often wide of the mark. Our perception of the sounds made by birds is often erroneous; one common fault, for example, is the difficulty that human hearing has with the rapid onset of many bird utterances that leads us to think we hear a starting consonant; in fact this is almost always untrue. Thus to convey correctly the sound of a call in words it is usually necessary to use consonants even though a bird sound usually does not have them. Generally the vowel sounds give fewer problems because they are longer. Phonetic renderings are highly subjective and descriptions of the same sound by different field workers often vary. Rarely are one’s expectations from reading a phonetic rendering matched in the field. However, many bird-watchers and ornithologists who are unfamiliar with sonagrams prefer using phonetic renderings to gain an insight into the type of sounds a bird makes. Indeed, such renderings are often simplified to mnemonics.
We usually try to describe sounds using phonetic renderings. However, we have made no attempt to standardize these renderings and have relied upon published sources or information supplied by our contributors, though we have, when possible, tried to indicate whether published examples are appropriate for the known vocalizations.
SONAGRAMS
Sonagram is the trade name for prints from a Kay Elemetrics Corp. ‘Sona-Graph’. The sound spectrograph or audio-spectrograph is an instrument that produces a frequency-spectrum-analysis. Most simply, sonagrams are graphs of frequency (pitch) against time (Fig. 1). Sonagrams are a permanent visual record of sounds. Sonagrams show, for example, the structure of a call or song, its duration, rate of repetition, intervals between utterances or the parts of utterances and the sound-frequencies involved. All sound is created by vibration; some vibrations are too slow and others too fast for the human ear to detect. Sound travels in waves and frequency is a measure of vibrations in these waves. Sounds of low frequency have long wave-lengths; sounds of high frequency have short wavelengths. The frequency of a sound is the main factor that determines the subjective pitch (which is the auditory perception of frequency); however, pitch varies with time, intensity and complexity of the sound (Hinde 1969).
Frequency is generally measured in cycles per second, the unit being a Hertz (Hz); scale is often given in kiloHertz (kHz). The higher the number, the higher the frequency or pitch of the sound and, on the sonagram, the higher above the baseline will be the trace. In conventional sonagrams, the frequency scale is portrayed as linear. Harmonics are exact doubling of frequency so that the musical notation of octaves expands through the range thus: 1–2 kHz is an octave; 2–4 kHz an octave and 4–8 kHz an octave. About five octaves fall below about 1 kHz but discrimination to the human ear is poor in the lowest part of this range. Most bird calls fall within the range of 1–8 kHz and are readily heard by the human ear. As a guide, the keyboard of the piano ranges from 27.5 Hz at bottom A to 4186 Hz at top C, with middle C at 261.63 Hz, and the human voice has peak energy levels (relative power) at about 400-600 Hz in men and 800-1000 Hz in women (Tremaine 1959). The smallest interval perceived by the human ear can be as little as 2 Hz. A semitone (the smallest musical interval in western music — the 12 keys on the piano through one octave), is about 60 Hz at the frequency of 1 kHz and therefore is easily measured in that part of the range or above on the sonagram. Many sounds have simultaneous emphasis of several frequencies; very often these frequencies are related as harmonics and these harmonics are the components of a sound wave that has frequencies that are simple multiples of the fundamental frequency. The fundamental frequency is the lowest component frequency of a complex sound wave. Amplitude is a measure of changes in air pressure of a sound; in a wave, it is measured as the maximum displacement of the wave. Amplitude is measured in decibels (dB), which is a relative term, but we give no precise measurement of amplitude in this Volume and need not discuss this unit further. Sonagrams readily show that sounds differ or resemble each other and allow similarities in calls or songs of different species to be objectively compared. Sounds that are identical can be recognized. Changes in pitch are easily seen; a monotonic whistle will show as a horizontal line for however long the whistle is held; a sound falling in pitch slopes downward from left to right; a sound increasing in pitch slopes upwards from left to right. Sonagrams also show amplitude (loudness) of a sound by the darkness of the tracing on the print. Contrary to many expectations, few bird sounds occur at high frequency but energy levels are often very high at frequencies where the sensitivity of the human ear is falling. Time resolution of sounds by birds is thought to be much better than in humans but there is some controversy on this matter (see Dooling 1982). Sounds fuse at about 50 ms intervals or less to the human ear, but it has been suggested that birds may be able to discriminate sounds only 5 ms apart.
RECORDINGS
Rather few studies of vocalizations of birds have been attempted in A’asia but there has been much interest in sound recording. At present, in Aust., recordings of all indigenous birds are being compiled and published (Buckingham & Jackson 1983, 1985, 1987, 1988, 1990, and in progress) and similarly for NZ (McPherson 1988, 1989a,b,c, and in progress). Fullagar & Robinson (1987) provide a guide to published discs and cassettes (likely to be available) and other sources of recordings of Aust. birds. Recordings of most Aust. and many New Guinean, NZ and Antarctic species are lodged with the ANWC — represented by the CSIRO Collection of Wildlife Sounds held at CSIRO Div. Wildl. Ecol., Canberra. In NZ, many recordings are held by NZ Wildl. Serv. Sound Library, NZDOC, Wellington. For more details on these and other major libraries of wildlife sounds see Kettle (1989).
The diversity of research on vocalizations of birds has resulted in many general reviews of the topic; important references are: Thorpe (1961), Hinde (1969), Thielcke (1976), Jellis (1977), Catchpole (1979), Kroodsma & Miller (1982) and Baker & Cunningham (1985) for dialects.
ARRANGEMENT
The Voice section is organized in two main parts. The first paragraph provides a general introduction to the species: any detailed studies (those that analyse voice and particularly include sonagrams); the main source(s) of information; how noisy or quiet the species is; general quality of vocalizations; periods of calling — daily, seasonally and annually; locations of calling (e.g. at nest-sites, over colonies). Individual and sexual differences and geographical variation in vocalizations are also discussed here. Non-vocal sounds are noted. Generally, we discuss all variation in sounds, regardless of scale over which that variation occurs, including dialects or geographical variation. Dialects are discussed only when they have been specifically discussed in a paper. The concept of dialects in vocalizations of birds is still controversial (see Baker & Cunningham [1985] and commentary and references therein for a full discussion). Dialects are a discrete and well-defined ‘population’ of vocalizations (e.g. song-types) shared by a local population of birds or subunits of that population. They occur when a population of birds uses a similar call- or song-type (or repertoire of types) different and discrete from neighbouring populations. Non-vocal sounds are summarized at the end of the first paragraph. The rest of the section deals with vocalizations and other sounds of adults and young. The descriptions are split into separate paragraphs for male and female if sexes differ. The last paragraph describes vocalizations of chicks and juveniles and, where known, the development of adult vocalizations. For reviews of the ontogeny of bird song see Konishi (1985).
Sonagrams have been included when recordings were available. Recordings from published sources have been preferred (specially those included by Buckingham & Jackson 1983, 1985 and McPherson 1988, 1989 a,b; mentioned above). Other recordings were sought only when these first sources did not include the required sound. Reference to the actual sound used for many sonagrams is therefore possible and the sound can be checked. Several other published recordings were needed to cover the full range of species included in this Volume and the full list is as follows: Bigwood & Bigwood (no date), Wildlife Service (1980), Scott 81. Dudderidge (1982), Crouch (1982), Griffin & Swaby (no date), Hutchinson (1972, 1978) and Gillard (1988). For some species published sound recordings were not available and recordings were sought from various sources. The British Library of Wildlife Sounds, London, kindly supplied material. Recordings held by The Library of Natural Sounds, Cornell Laboratory of Ornithology, Ithaca, New York, the CSIRO Collection of Wildlife Sounds, Canberra and some uncatalogued material from private sources were needed to complete our coverage. Many sounds could not be illustrated because we could not trace an appropriate recording and most species accounts fall far short of being comprehensive; for most species we can do no more than present some of the more common and well-known vocalizations or other sounds as sonagrams. Captions to each sonagram indicate details of source, date and locality of recording and recorder. All recordings were analysed in the sound laboratory of the CSIRO Division of Wildlife and Ecology using a Kay Elemetrics Corp. 7800 digital storage Sona-Graph and a Kay 7900 hard-copy printer. We are specially indebted to Sir Frederick White FRS, former Chairman of CSIRO, who generously made possible the purchase of this equipment with the intention that it be used in part for the HANZAB project. All sonagrams are reproduced direct from the original prints to retain the true tonal contrasts but, as far as possible, irrelevant sounds (background noises and intrusions) have been eliminated. All recordings were analysed using 0–8 kHz scale with a 300 Hz analysis filter. Each sonagram is shown with a contrasting overlay (red) giving scale for time divisions at 100 ms and frequency divisions at 1 kHz.
REFERENCES
- Armstrong, E.A. 1969. Pp. 343-65. In: Hinde 1969.
- Baker, M.C., & M.A. Cunningham. 1985. Behav. Brain Sci. 8: 85- 133.
- Bennet-Clark, H.C. 1981. In: McFarland 1981.
- Bigwood, K., & J. Bigwood. N.d. A Treasury of New Zealand Bird Song. (Cassette). Kiwi Record TC SLD-25.
- Brackenbury, J.H. 1982. Pp. 53-73. In: Kroodsma & Miller 1982.
- Brockway, B.F. 1962. Living Bird 1: 93-101.
- Brockway, B.F. 1965. Anim. Behav. 13: 575-78.
- Buckingham, R., & L. Jackson. 1983. A Field Guide to Australian Bird Song. Cassette 1. Bird Obs. Club.
- Buckingham, R., St L. Jackson. 1985. A Field Guide to Australian Bird Song. Cassette 2. Bird Obs. Club.
- Buckingham, R., & L. Jackson. 1987. A Field Guide to Australian Bird Song. Cassette 3. Bird Obs. Club.
- Buckingham, R., & L. Jackson. 1988. A Field Guide to Australian Bird Song. Cassette 4. Bird Obs. Club.
- Buckingham, R., & L. Jackson. 1990. A Field Guide to Australian Bird Song. Cassette 5. Bird Obs. Club.
- Campbell, B., & E. Lack. (Eds) 1985. A Dictionary of Birds.
- Catchpole, C.K. 1979. Vocal Communication in Birds.
- Catchpole, C.K. 1982. Pp. 297-319. In: Kroodsma & Miller 1982.
- Crouch, H. N.d. Birds Calls of Rotamah Island. (Cassette).
- Dooling, R.J. 1982. Pp. 95-130. In: Kroodsma St Miller 1982.
- Fletcher, N.H. 1988. J. Theor. Biol. 135: 455-81.
- Fullagar, P.J., & F.N. Robinson (Eds) 1987. A Guide to Recorded Australian Wildlife Sounds: Birds. AGPS.
- Gaunt, AS. 1983. Auk 100: 853-62.
- Gaunt, A.S., & S.L.L. Gaunt. 1985. Current Ornithology 2: 213- 45.
- Gillard, L. 1988. Bird Calls of Eastern Australia. (Cassette).
- Greenewalt, C.H. 1968. Bird Song: Acoustics and Physiology.
- Griffin, A.A., & R.J. Swaby. N.d. Birds Calls of Tropical Eastern Australia. (Cassette).
- Hinde, R.A. (Ed.) 1969. Bird Vocalizations.
- Hutchinson, J.N. 1972. Australian Bird Calls Index: Ser. 1. Western Australia. (Cassette).
- Hutchinson, J.N. 1978. Australian Bird Calls: Ser. 2. Western Australia. (Cassette).
- Jellis, R. 1977. Bird Sounds and their Meaning.
- Kettle, R. 1989. Bioacoustics 2: 171-6.
- Konishi, M. 1985. Ann. Rev. Neurosci. 8: 125-70.
- Kroodsma, D.E., & E.H. Miller. (Eds) 1982. Acoustic Communication in Birds.
- McFarland, D. (Ed.) 1981. The Oxford Companion to Animal Behaviour.
- McPherson, L.B. 1988. New Zealand Birds: A Sound Guide. Cassette 1
- McPherson, L.B. 1989a. New Zealand Birds: A Sound Guide. Cassette 2
- McPherson, L.B. 1989b. New Zealand Birds: A Sound Guide. Cassette 3
- McPherson, L.B. 1989c. New Zealand Birds: A Sound Guide. Cassette 4.
- Nowicki, S. 1987. Nature 325: 53-5.
- Scott, P., & P. Dudderidge. 1962. Wildfowl Calling. (Record) EMI Records.
- Thielcke, G.A. 1976. Bird Sounds.
- Thorpe, W.H. 1961. Bird Song.
- Tremaine, H.M. 1959. The Audio Cyclopedia.
- Warner, R.W. 1972. J. Zool., Lond. 168: 381-93.
- Wildlife Service. 1980. Birds of New Zealand: 38. Forest and Sea Birds. (Cassette) VP445C. NZ Dept Int. Affairs.
From Vol. 2
Sonagrams are included where suitable recordings were available. Recordings were analysed in the sound laboratory of the Australian National Wildlife Collection using a Kay Elemetrics Corp 7800 sonagraph and a Kay 7900 printer. Recordings were analysed over 0-8 kHz using a 300 Hz analysis filter. Each sonagram is shown with an overlay with time-scale (x-axis) divisions every 0.5 s and frequency scale divisions (y-axis) every 1 kHz. The amplitude (loudness) of a sound is shown by the darkness of the tracing on the print. To retain tonal contrasts, sonagrams are reproduced directly from the original prints but irrelevant intrusions and background noises have, as far as possible, been eliminated.
To permit reference to the actual sound used to make a sonagram, published recordings have been used as much as possible, particularly those from the compilations attempting to cover all species in our region (Buckingham & Jackson in Aust.; McPherson for NZ). Recordings from the sound library of the ANWC have been used to complete the coverage. The caption to each sonagram lists the recordist, place and date of the recording, and the source of a recording. The source is a letter followed by a number; the letter P indicates a published recording. The published recordings used in Volumes 1 and 2 of HANZAB are listed below.
In this Volume, the survey and review of published information on calls of shorebirds by Miller ( 1992) was a valuable reference.
Published sound recordings
- P10 Hutchinson, J.N. 1972. Australian Bird Calls, Ser. 1. Author, Balingup, WA.
- P11 —1978. Australian Bird Calls, Ser. 2. Author, Balingup, WA. [added Vol. 5]
- P19 Crouch, H., & A. Crouch. 1982. Birds of Rotamah Island. Authors, Belair, SA.
- P26 Buckingham, R., & L. Jackson. 1983. Field Guide to Australian Birdsong. 1: Emu to Striated Heron. BOCA, Melbourne.
- P28 van Gessel, F.W. No date [c. 1984]. Bird Calls of the Hunter Region. 1 (Tape 1). Bushbirds. Author, Aust. [added Vol. 5]
- P30 Flentje, W.M. 1980. Some Birds of the Victorian Forests. Author, Bendigo, Vic.
- P32 Hutchinson, J.N. No date. Australian Bird Calls, Ser. 4. Author, Balingup, WA.
- P34 Crouch, H., & A. Crouch. 1982. Bird Calls of Vicroria: Urban Areas. Authors, Belair, SA.
- P35 Buckingham, R., & L. Jackson. 1985. Field Guide to Australian Birdsong. 2: Rufous Night Heron to Chestnut Rail. BOCA, Melbourne.
- P36 —, —,1987. Field Guide to Australian Birdsong. 3: Red-necked Crake to Crested Tern. BOCA, Melbourne.
- P37 Griffin, A., & R.J. Swaby. No date. Bird Calls of Tropical Eastern Australia. Authors, Paluma, Qld.
- P38 van Gessel, F.W. No date [c. 1987]. Bird Calls of the Hunter Region. 1 (Tape 2). Honeyeaters. Author, Aust. [added Vol. 5]
- P39 Buckingham, R., & L. Jackson. 1988. A Field Guide to Australian Birdsong. 4: Sooty Tern to Superb Parrot. Birds Obs. Club Aust., Melbourne. [added Vol 2]
- P40 —, — 1990. A Field Guide to Australian Birdsong. 5: Regent Parrot to Masked Owl. Bird Obs. Club Aust., Melbourne. [added Vol. 4]
- P43 Gillard, L. 1988. Bird Calls of Eastern Australia.
- P48 Buckingham, R., & L. Jackson. 1991. A Field Guide to Australian Birdsong. 6: Eastern Grass Owl to Ground Cuckoo-shrike. Bird Obs. Club Aust., Melbourne. [added Vol. 4]
- P49 Buckingham, R., & L. Jackson. 1992a. A Field Guide to Australian Birdsong. 7. White-winged Triller to White-breasted Whistler. Bird Obs. Club Aust., Melbourne. [added Vol. 6]
- P50 —, — 1992b. A Field Guide to Australian Birdsong. 8. Little Shrike-thrush to Hall’s Babbler. Bird Obs. Club Aust., Melbourne. [added Vol 6]
- P63 Flentje, W.J. 1992. Night Sounds of the Forest. Author, Bendigo, Vic. [added Vol. 4]
- P73 Buckingham, R., & L. Jackson. 1994. A Field Guide to Australian Birdsong. 9. Chestnut- crowned Babbler to Redthroat. Bird Obs. Club Aust. , Melbourne. [added Vol. 5]
- P75 —, — 1995. A Field Guide to Australian Birdsong. 10. Calamanthus to Noisy Friarbird. Bird Obs. Club Aust., Melbourne. [added Vol. 5]
- P78 McNabb, E.G. 1995. Nightlife of the Dandenongs. Author, Emerald, Vic. [added Vol. 4]
- P90 Buckingham, R., & L. Jackson. 1997. A Field Guide to Australian Birdsong. 11. Little Friarbird to Scarlet Honeyeater. Bird Obs. Club Aust., Melbourne.
- P94 —, — 1999. A Field Guide to Australian Birdsong. 12. Crimson Chat to Torresian Crow and supplementary calls. Bird Obs. Club Aust., Melbourne.
- P100 Wildlife Service, NZ Dept Int. Afr. 1980. Birds of New Zealand: 38 Forest and Sea Birds. Viking Music Cassette VP445C.
- P101 Scott, P, & P Duddridge. 1962. Wildfowl Calling. EMI Records, Hayes, Middlesex, UK.
- P102 Bigwood, K., & J. Bigwood. No date. Treasury of New Zealanel
- Bird Song. Kiwi Record TC SLD-25.
- P103 McPherson, L.B. 1989. New Zealand Birds: A Sound Guide 3. Author, Christchurch.
- P104 — 1989. New Zealand Birds: A Sound Guide. 4. Author, Christchurch.
- P105 — 1990. New Zealand Birds: A Sound Guide. 5. Author, Christchurch.
- P106 — 1990. New Zealand Birds: A Sound Guide. 6. Author, Christchurch.
- P107 — 1990. New Zealand Birds: A Sound Guide. 7. Author, Christchurch.
- P108 Chappuis, C. No date. Sounds of Migrants and Wintering Birds
- in Western Europe. Vol. 2.
- Q2 Ulman, P., & J. Ulman 1991. Mountain Scenes, Nature’s Sounds, Aust. Broadcasting Corporation, Sydney.
- Q29 Horton, H. 1996. Birdsong of Australia: Birdcalls of the Desert and Semi-arid Regions of Inland Australia. Aust. Broadcasting Corporation, Sydney.
- Q99 Stewart, D.A. 2002 Australian Bird Calls: Tropical North-east. Nature Sound, Mullumbimby, NSW.
REFERENCES
- Miller, E.H. 1992. Acoustic Signals of Shorebirds. Royal Br. Columbia Mus. Tech. Rep., Victoria, Be, Canada.
From Vol. 3
Sonagrams are included where suitable recordings were available. Recordings were analysed using Apple Macintosh computers. Sounds were digitized at 16-bit resolution and edited using SoundEdit software (Macromedia Inc.). Sonagrams were made using Canary 1.1 software (Bioacoustics Res. Prog., Cornell Lab. Orn.). Sonagrams were sent in electronic form (as PICT files) to the publisher. Recordings were analysed over a frequency range of 0-8 kHz using a Hamming window function, a filter bandwidth of c. 350 Hz, smooth display style, 50% overlap and 256 point FFT size. Each sonagram is shown with an overlay with time on the horizontal scale and frequency on the vertical scale. The amplitude (loudness) of a sound is shown by the darkness of the tracing. Irrelevant
intrusions and background noises have, as far as possible, been removed.
[Rest of text as per text for Vol 2 except for the addition of the inclusion of P39 in the list of published sound recordings which has been added to the list for Vol 2]
From Vol. 4
Sonagrams were made using Canary 1.2 software (Bioacoustics Res. Prog., Cornell Lab. Orn.) on Apple Macintosh computers. Sounds were digitized at 16-bit resolution and edited using SoundEdit software (Macromedia Inc.). Irrelevant intrusions and background noises were, as far as possible, removed. The analysis used a Hamming window function, a filter bandwidth of c. 350 Hz, smooth display style, 50% overlap and 256 point FIT size. Sonagrams were sent in electronic form (as PICT files) to the publisher. Each sonagram is shown with an overlay, with time on the horizontal scale and frequency on the vertical scale. The amplitude (loudness) of a sound is shown by the darkness of the tracing.
Because of the need to analyze calls longer than those shown hitherto, we have abandoned the fixed time-scale used previously. Sonagrams one column wide may show 2.5 s, 5 s, 10 s or 20 s of sound. The vertical frequency scale of 0–8 kHz has, by and large, been retained, regardless of the length of time shown, so that slopes of ascending and descending calls will appear steeper in sonagrams showing longer durations of sound.
In a few sonagrams the vertical (frequency) scale extends beyond the usual 8 kHz.
As in earlier volumes, to permit reference to the actual sound used to make a sonagram, published recordings have been used as much as possible. Recordings from the sound library of the Australian National Wildlife Collection (ANWC), CSIRO Division of Wildlife and Ecology, Canberra, or from private collections have been used to complete the coverage; in particular we have made extensive use of the large collection (c. 140 h of edited calls on digital tape) recently contributed to the ANWC by D.A. Stewart. The caption to each sonagram lists the recordist, place and date of the recording, and the source of the recording. If the source of the recording is preceded by letter P, then it is a published recording and is listed [under Volume 2].
From Vol. 5
There have been no changes to the scope or arrangement of this section.
For this volume, sonagrams were made, where suitable recordings were available, using an Apple Macintosh computer. Sounds were digitized at 16-bit resolution and edited using SoundEdit software (Macromedia Inc. ). Irrelevant intrusions and background noises were, as far as possible, removed. Sonagrams were made using Canary 1.2.4 software (Bioacoustics
Res. Prog., Cornell Lab. Om.). The analysis used a Hamming window function, a filter bandwidth of c. 350 Hz, smooth display style, 50% overlap and 256 point FFT size. Sonagrams were sent in electronic form (as PICT files) to the publisher. Each sonagram is shown with an overlay, with time on the horizontal scale and frequency on the vertical scale. The amplitude (loudness) of a sound is shown by the darkness of the tracing.
To permit reference to the actual sound used to make a sonagram, published recordings have been used as much as possible, particularly those from the compilations attempting to cover all species in the HANZAB region (Buckingham & Jackson for Aust.; McPherson for NZ). Recordings from the sound library of the Australian National Wildlife Collection, CSIRO Division of Wildlife and Ecology, Canberra, have been used to complete the coverage; in particular we have made much use of the large collection contributed by D.A. Stewart. The caption to each sonagram lists the recordist, place and date of the recording, and the source of the recording. If the source is a number prefixed by the letter ‘P’ then the source is a
published recording and is listed [see list under Vol 2 above].
Sonagrams show 2.5 s, 5 s, or 10 s of sound; there is also one sonagram that shows 198 s of the Sequential Song of the Albert Lyrebird. The vertical frequency scale does not change, regardless of the length of time shown, so that slopes of ascending and descending calls will appear steeper in sonagrams showing shorter durations of sound. Reader beware! For species with calls of very high frequency (e.g. for the Acanthisittidae, Maluridae, and Stitchbird Notiomystis cineca) the vertical (frequency) scale extends to 16 kHz.
We wish to pay tribute to the completion of A Field Guide to Australian Birdsong, a major project on the vocalizations of Aust. birds, to which many sound recordists have contributed, and which was compiled over two decades by R. Buckingham and L. Jackson for the Bird Observers Club of Australia. Unfortunately R. Buckingham died in July 1999, a few months before publication of the last of the 12 cassettes. The series has been of great value in preparing all volumes to date, and its sounds have been the source of most of the sonagrams.
From Vol. 6
[Vol 7 similar and not repeated]
Sonagrams were made, where suitable recordings were available, using an Apple Macintosh computer. Sounds were digitized at 16-bit resolution and edited using SoundEdit software (Macromedia Inc.). Irrelevant intrusions and background noises were, as far as possible, removed. Sonagrams were made using Canary 1.2.4 software (Bioacoustics Res. Prog., Cornell Lab. Orn.). The analysis used a Hamming window function, a filter bandwidth of c. 350 Hz, smooth display style, 50% overlap and 256 point FFT size. Sonagrams were sent in electronic form (as PICT files) to the publisher. Each sonagram is shown with an overlay, with time on the horizontal scale and frequency on the vertical scale. The amplitude (loudness) of a sound is shown by the darkness of the tracing.
To permit reference to the actual sound used to make a sonagram, published recordings have been used as much as possible, particularly those from the compilations attempting to cover all species in our region (Buckingham & Jackson for Aust.; McPherson for NZ). Recordings from the sound library of the Australian National Wildlife Collection (CSIRO Division of Sustainable Ecosystems, Canberra) have been used to complete the coverage; we have also made use of the large collection contributed by D.A. Stewart. The caption to each sonagram lists the recordist, place and date of the recording, and the source of the recording. If the source is given as the letter P followed by a number then the source is a published recording and is listed [see list under Vol 2 above]. From Vol 7 – … If the source is given as the letter P (records, cassettes] or Q (compact discs) …
Sonagrams show 2.5 s, 5 s, or 10 s of sound. The vertical frequency scale is used regardless of the length of time shown, so that increases or decreases in pitch appear to be more rapid in sonagrams showing longer durations of sound. Reader beware! For sonagrams of the White-browed Scrubwren Sericomis frontalis the vertical (frequency) scale has been extended, and for one sonagram for this species a different horizontal scale has been used.
In the introduction to Volume 5 we paid tribute to the completion, in 12 cassettes, of A Field Guide to Australian Birdsong, compiled for the Bird Observers Club by the late R. Buckingham and by L. Jackson. This compilation has continued to be of great value, and its sounds have been the source of most sonagrams. The Bird Observers Club has begun a revised edition on compact disc, compiled and edited by H. Plowright, and intended for quick reference. It does not replace the cassette edition with its comprehensive booklets, and a particular track from the cassette edition may be abbreviated or omitted in the CD edition, which also contains
some new recordings. The sonagrams in this volume that are made from calls in A Field Guide to Australian Birdsong are from calls in the cassette edition, which mayor may not be included in the CD edition.
BREEDING
[Information from Vol. 1, 2, 3, 4, 5, 7]
From Vol. 1
This section starts with a paragraph that briefly outlines the chief sources of information, mentions the contributor of the information and summarizes the sort of breeding habits and habitat of the species. The chief references are identified with the place at which the work was done. Thereafter in the text, reference is made only to the place where the observations were made and it can be assumed that the reference is to the publication already mentioned for that place. In this way, we avoid a litany of bracketed references throughout, which can only irk a reader. The section is divided into subsections for Breeding Season, Nest-site, Nest and Materials, Eggs, Clutch-size, Laying, Incubation, Young (Nestling), Growth, Fledging to Maturity and Breeding Success.
SEASON
We take the View that breeding seasons can be determined only by reference to the date of laying of the first egg in a clutch. Pre-laying activity that heralds nesting, such as displays, pair-formation and the like, cannot be timed so exactly and varies in duration. Once a clutch is started, all other breeding events (hatching, fledging, etc) follow inevitably and their timing can be calculated if the periods of incubation and fledging are known. Moreover, once a bird starts to lay, it will go on doing so with later broods or with replacement clutches after loss until something intervenes to stop it, such as regression of the gonads, moult or even inclement weather. Those birds that lay only one egg in a season and make no attempt to replace it after loss or to start a second brood after success are merely the start of a line that ends with species that have multiple broods or will replace losses many times. It is less significant to record young in a nest than to be able to estimate within reasonable limits when the eggs that produced those young were laid. In the accompanying diagrams of annual breeding cycles (see below), we have therefore tried to emphasize the period of laying. These diagrams are in linear form and not in the customary circular form because the linear form saves space, allows the depiction of events at more than one place and is the only convenient way of displaying the breeding cycle of species that breed only in every second year. The breeding season is so sketchily known for some species that a diagram is not worthwhile; it could only mislead. Most pelagic species such as penguins and procellariiforms have synchronized periods of laying and their annual cycle is well adapted for the linear form of diagram where also periods of hatching, creching, fledging and moult can readily be shown to help people to see quickly when the birds are likely to be at different stages in the reproductive cycle. On the other hand, diagrams are difficult to draw for many species of waterbirds in Aust. For various reasons, the nests of some species of cormorants, herons, ibis, ducks and others are hard to watch systematically, and in consequence, exact data for many aspects of their nesting are few. Such as there are generally provide little information beyond occupation of active nests and have been reported in such a way that ageing of the nest is mere guesswork. Yet, it seems that for many widely distributed species, populations in n. Aust. lay mostly in autumn to spring (Apr.–Aug.) whereas those in s., se. and sw. Aust. lay mostly in spring and summer (Sept.–Jan.). Undoubtedly, many of these species have protracted laying periods but for all or most of them the matter is greatly complicated by the influence of erratically fluctuating wet and dry conditions that cannot be predicted. Thus, what happens in a locality in one season is a poor guide to what will happen in the next. The facts, such as we have judged them to be, are given in the text, supplemented by a diagram where it seems justified, but there is still much scope for observers to correct and improve our interpretation. Diagrams of breeding cycles. For many pelagic and colonial species (e.g. Fig. 1), we show dates of arrival, laying, hatching, departure and other events, such as formation of creches, fledging (before departure) and independence of young. For other species, mostly land birds (e.g. Fig. 2), we simply show when eggs are laid, by a solid black bar, which may taper to show increases or decreases in laying; black dashes indicate when eggs have been found. When available, information on primary moult is shown in the narrow bar below.

NEST-SITE, NEST AND MATERIALS
Most species dealt with in Volume 1 do not place their nests in obviously special sites, build rather simple nests, if any at all, and for the most part use any material that is at hand. Measurements of all sorts relating to aspects of sites, height above ground, depth of water, size and shape of nest and so on are mostly imprecise. In consequence, information on these matters is usually generalized. The subject could as well be treated under one heading and usually needs further investigation. However, we have tried to make a distinction between the precise siting of a nest and the nest itself, even if that is not clear in, say, species that nest in burrows. Broader aspects of nesting habitat are given under Habitat. Material used in nests is not often described in detail in the literature and often we can only follow with generalizations. The roles of the sexes in selection of sites and building, and the pattern of building, are also not well recorded and often need further investigation. EGGS. The shape, texture and colour of eggs are described briefly, mostly after descriptions by earlier authors like Campbell and North. Measurements are always in millimetres arranged as average length (standard deviation, if available; range; size of sample) x average breadth (SD; range). The size of the sample thus ranges from satisfactorily large (>50) to very meagre (<10) or even to measurements of single eggs. Where possible, measurements from different parts of the breeding range are given. Because so many different species in this Volume lay uniformly white or pale-coloured eggs, descriptions even with illustrations do not help identification greatly and measurements are more important in this respect. Schönwetter’s work (1967, et seq.) has been used for some species but we have not incorporated it for others that have been studied well by other workers.
CLUTCH-SIZE
As far as possible, data are presented only from nests in which laying has been recorded to completion (i.e. from the first egg to the same number of eggs for two inspections made at an interval greater than the normal interval of laying or for those with the same number of eggs on two inspections during incubation at an interval greater than that of laying). The data are given as an average (standard deviation, if available; range; number of nests). If such quantified data are not available, and even sometimes as a supplement to small quantified samples, the range as generalized in standard references is stated, with the qualifying phrase ‘said (claimed) to be …’ to show the unsatisfactory state of our knowledge. For those species that lay few eggs (1-2), there are no problems except perhaps for the controversial finding of three eggs in the nests of some penguins. For species with large clutches, there are problems. Nests of colonial species may be difficult to visit with the regularity needed for the accurate determination of completed clutches, either because visits to a colony cannot or have not been made often enough or because nests are not readily accessible. Thus the reliability even of quantified data may be doubtful. Further, there is often a possibility or probability of two females laying in the same nest and that leads on to the supreme difficulty of dump-nesting in ducks. If a respectable sample of critically assessed clutches is not available for a quantified presentation, the few recorded acceptable clutches are given in detail. Where dump-nesting is proved or suspected, there is little to be done beyond mentioning the likely limits of true clutches. Sometimes it has to be admitted that we do not know what the clutch-size of an individual may be. For many species, there is need for more critical observation of clutch-size. Replacement of clutches in a season is discussed in this subsection.
LAYING
Primarily this part is for recording the intervals at which, and the time of day when, birds lay their eggs in a clutch, again a matter that is usually poorly known. Details of synchronization of laying in colonially nesting species, which may have been mentioned in the subsection for Season, are given here.
INCUBATION
The start of incubation, the role of the sexes and the pattern of shifts of incubation are given. For some closely studied species, especially among penguins and petrels, these can be given in considerable detail but in other groups there is often little information. Though these aspects are generally similar among all members of a family or genus and may be inferred without much danger of error, descriptions are given only where the behaviour has actually been recorded. The incubation period is detailed, where possible, as the period from laying to hatching of individual eggs or as the period from laying of the last egg of a clutch to the hatching of the last young. Most of the species covered in Volume 1 have long periods (>25 days) and more variation occurs than in birds like passerines with short periods (<20 days). Moreover, especially among the procellariiforms, eggs may survive quite long periods of desertion. In consequence, there is not much object in trying to express the incubation period within narrow limits because that could give an impression only of spurious accuracy. All the same, more investigation and recording of the period is needed for many species.
YOUNG OR NESTLING
Either term may be used because, though a young petrel that stays in its burrow until fledging may reasonably be called a nestling, a duckling obviously cannot be so described. The state of development at hatching is given first in standard terms (altricial, semi-altricial or precocial) with those for subsequent behaviour (nidicolous, semi-nidicolous or nidifugous) being generally characteristic of whole orders or families. Some description of the young and its plumage development is then given (fuller details in Plumage section), designed, it is hoped, to enable observers to give an approximate age to young seen on a single visit and thus be able to estimate a date for laying of the clutch, which greatly enhances the value of a record. Parental care and the roles of the sexes in brooding, guarding and feeding the young are then described but only when they have been definitely recorded for a species. These activities, being characteristic of orders and families rather than of species, can be inferred to occur in all members of the larger groups but we mention them only if that has been confirmed and recorded. In many species covered in Volume 1, parents feed their young by regurgitation: complete if the food is spewed out onto nest or ground; incomplete if the food is retained in the parent’s beak or throat and collected there by the young. The nestling or fledging period of species in Volume 1 is usually poorly known and often varies quite considerably, being influenced by climatic factors, an irregular food supply or other variables. Moreover, the age at departure from a colony or the first flight of precocial, nidifugous or semi-nidifugous young may be hard to determine. In any case, this period usually lasts for several or many weeks and it is hardly important to determine it closely.
GROWTH
Some details of growth (weights, measurements, etc.) may be given here, as further help towards ageing of young in the field; references are made to original studies or to the Plumage section for full details.
FLEDGING TO MATURITY
The period during which young depend on their parents after fledging or first flight is mostly known very poorly. It may be perfectly correct to assume that the young of pelagic species are completely independent once they go to sea, but for other species there is little known about the matter. Similarly, the ages at which birds first return to colonies, form pairs and start to breed are known in only a few species.
BREEDING SUCCESS
Hatching, fledging and total success are given, even if samples are small and apply only to a single season. They may have little significance for long-lived species in which reliable figures can be had only from long-term studies, especially in erratically fluctuating conditions such as prevail in Aust. Here mention is made of what little is known of predation and loss of eggs and young, mostly greatly generalized.
REFERENCES
- Schönwetter, M. 1967, et seq. Handbuch der Oologie.
From Vol. 2
Season
For many terrestrial species in Aust. and NZ, knowledge of breeding is often so poor that it is doubtful whether generalized linear diagrams for the breeding (laying) season arc useful or practical. We have used such diagrams sparingly.
From Vol. 3
Arrangement follows that of Volumes 1 and 2. In this Volume, we have not used generalized linear diagrams to show the breeding season.
From Vol. 4
Remains much as previous volumes, with no significant changes.
FLEDGING and FLEDGING PERIOD
There is much confusion in the literature concerning the term ‘fledging’. Some authors define it as when a bird first leaves the nest; others, when a bird leaves the nest permanently; others still, when a bird first flies. Often, what individual authors intend or mean is not at all clear. We define fledging as when a young bird first leaves the nest; a fledgeling as a young bird that has left the nest at least once; and fledging period as the time from hatching till a bird first leaves the nest. All definitions have to deal with the problem of premature fledging, such as in alarm or other circumstances, or when a bird leaves the nest, often for increasing periods of time, but returns to it in between. The advantage of the definitions adopted is that it is not necessary to determine subjectively what the first flight may be, nor determine what constitutes permanent departure from the nest.
The paragraph, Fledging to maturity, has been changed to Fledging to independence, because maturity is not easily defined. Independence, as we use the term, refers to the stage at which a fledgeling is no longer dependent on its parents or parent for food or protection or both. Age of first breeding is now given in Social Organization (in Bonds).
From Vol. 5
Arrangement essentially follows that of previous volumes with no major changes. There are a few points we wish to reiterate.
Young
In this paragraph, we use the following terms to describe the state of young at hatching (altricial, semi-altricial or precocial) and subsequent behaviour (nidicolous, seminidicolous or nidifugous) (from Campbell & Lack 1985 ):
ALTRICIAL: Hatch with little or no down, with eyes closed; young unable to leave nest and are fed by parents.
SEMI-ALTRICIAL: At hatching, covered in down and eyes are open or closed; young unable to leave nest and are fed by parents.
PRECOCIAL: Hatch covered in down and with eyes open; young leave nest on first or second day and usually follow parents but find own food.
SEMI-PRECOCIAL: Hatch covered in down and with eyes open; young unable to leave nest and are fed by parents.
NIDICOLOUS: Young remain in nest after hatching.
NIDIFUGOUS: Young leave nest immediately, or soon after, hatching.
Fledging to independence
This paragraph was previously labelled Fledging to maturity but was changed from Volume 4 on because maturity is not easily defined. Independence, as we use the term, refers to the stage at which a fledgeling is no longer dependent on its parents or parent for food or protection or both and, while not always exact, is often better known than time of maturity. Age of first breed ing is now given in Social Organization (in Bonds).
FLEDGING and FLEDGING PERIOD
There is much confusion in the literature concerning the term ‘fledging’. Some authors define it as when a birds first leaves the nest; others, when a bird leaves the nest permanently; others still, when a birds first flies. Often, what individual authors intend or mean is not at all clear. We define fledging as when a young bird first leaves the nest; a fledgeling as a young bird that has left the nest at least once; and fledging period as the time from hatching till a bird first leaves the nest. All definitions have to deal with the problem of premature fledging, such as in alarm or other circumstances or when a bird leaves the nest, often for increasing periods of time, but returns to it in between. The advantage of the definitions adopted is that it is not necessary to determine subjectively what the first flight may be, or determine what constitutes permanent departure from the nest.
Success
This was calculated for nests where clutch-size, number of eggs hatched and number of young fledged was certain. Nests that failed during laying were included in calculations of egg (hatching) success. Success was not calculated for nests where young were seen in the nest close to fledging but no fledged young could be found.
REFERENCES
- Campbell, B., & E. Lack. (Eds) J 985 . A Dictionary of Birds. T. & A.D. Poyser, Calton, England.
From Vol. 7
In our discussion of Season, we have sometimes included a monthly breakdown of breeding records from either the first Atlas of Australian Birds or the New Atlas of Australian Birds (Aust. Atlas 1, 2 [see Abbreviations and Conventions: Standard references]) or both combined. In these cases, the data comes from Atlas databases and not the published books.
PLUMAGES AND RELATED MATTERS
From Vol. 2
Conventions used are outlined fully in Volume 1 of HANZAB. For the most part these should be clear from the texts but some points merit reiteration or clarification.
From Vol. 7
These sections are primarily based on examination of material within the collections of museums in Aust., NZ and, less often, elsewhere, as well as a compilation of available literature. For each species, the editor or editors who undertook the primary research and prepared the sections on Plumages and related matters is named at the start of the Plumages section.
PLUMAGES
[Information from Vol. 1, 2, 3, and 5,6,7]
From Vol. 1
All feathered parts are described here; the rest of the integument is described in the section Bare Parts. The descriptions in this section are based on museum skins, unless otherwise stated. The section is intended chiefly for use with the bird in the hand, but may be helpful in other situations. Usually only one subspecies is described; others are covered in the section Geographical Variation. Plumages are dealt with in the sequence: adult breeding, adult non-breeding, nestling or downy young, juvenile and immatures. When plumages differ between the sexes, males are described first.
Nomenclature of Plumage and Moult
Two terminologies for moults and plumages are widely used. That introduced by Dwight (1900), with slight modifications, was used in BWP and is generally used in the Old World. The terminology introduced by Humphrey & Parkes (1959) has a strong following in North America. We have used the terminology of BWP for all plumages, because it is more familiar to A’asians and more readily applied to bare parts. Sometimes, this terminology may give a misleading impression of a plumage, and then we have also applied the nomenclature of Humphrey & Parkes, or given additional notes, or both. Accordingly, both schemes are summarized in Table 1 and outlined below.

A plumage is a single generation of feathers, which may not include the entire feather covering of a bird; feathers attained by a partial moult also constitute a plumage. Wear of feathers or transitional stages from one plumage to another often affect appearance. Where we have felt it necessary to cover such variation under separate headings we have done so.
In the BWP scheme, breeding plumage is defined as that worn during part or all of the breeding season. In some species, the breeding plumage alternates regularly with a non-breeding plumage, acquired during the post-breeding moult. In others, one plumage is worn throughout the cycle (defined in glossary) and is termed adult. The first pennaceous plumage of a bird is called juvenile in all species, even in those in which it appears identical to adult plumage. In some species, juvenile plumage is followed by one or more recognizable immature plumages. These are called first immature, second immature, etc. in increasing order of age. If there are no other recognizable plumages, the juvenile plumage is said to be replaced by the first adult plumage.
The above nomenclature for plumages is easily understood, and is also popular because it ‘links the various plumages with the phases in the life cycle for which they have evolved’ (BWP). However, there are examples in which the nomenclature is inappropriate. For example, the terms ‘adult’ and ‘immature’ suggest sexual maturity, or lack of it. This is clearly misleading in procellariiforms, most of which have an adult plumage for several years before they begin to breed. It is equally confusing in Golden Whistlers Pachycephala pectoralis; immature males can breed in a plumage that resembles the adult female rather than the adult male. In many species, the above terminology implies a relation between plumages and breeding or development that has not actually been determined. Where the nomenclature of BWP seems open to misinterpretation we have drawn attention to it next to the plumage headings.
The nomenclature for moults and plumages introduced by Humphrey & Parkes (1959) has a terminology independent of other phases in the life cycle. The scheme has also been described by Palmer (1962, 1972, 1988) and Wilds (1989). It is difficult to explain their scheme without mentioning their nomenclature for moult. They named moults by adding ‘pre-’ to the name of the plumage they produce.
Adult birds that have one plumage per cycle almost invariably lose and renew their plumage in a complete moult. This is called the basic plumage; it is renewed in the pre-basic moult. The pre-basic moult can also be recognized as a complete moult in adult birds that have two plumages per cycle. These birds have a separate moult, usually partial, termed the pre-alternate moult. It results in an alternate plumage; birds in alternate plumage generally also have retained feathers (usually remiges and rectrices) from the basic plumage. A few birds have more than two moults in a cycle. The plumage resulting from such an additional moult is called the supplemental plumage. It may be worn before or after the alternate plumage.
As in the BWP scheme, the first pennaceous plumage is termed juvenile. It can be replaced in a complete moult, but more commonly there are stages of partial moult. The moult in which replacement of juvenile plumage begins must be termed pre-basic, whether it is complete or not, because it is the only moult that can be expected in the first cycle of all birds. In species that have identifiable age-classes for some time, it is convenient to qualify the names of the plumage stages with a numerical prefix. This results in such terms as first basic plumage, first pre-basic moult, etc. Humphrey & Parkes suggested that the term definitive be applied to plumages that do not change any more with age.
Some aspects of Humphrey & Parkes nomenclature have been criticized (Stresemann 1963; Amadon 1966), particularly the underlying assumption that the basic plumage is homologous across groups of birds. Even if this assumption is untrue, the nomenclature often has advantages. Examples in which the terminology of BWP is misleading were mentioned above. There is no misinformation conveyed when an adult procellariiform is termed definitive basic, or an immature male Golden Whistler is said to be in its first basic plumage.
Humphrey & Parkes’ nomenclature has not been applied to all plumages in this book, because it is often difficult to use. Partial moults of body-feathers that produce no change in the appearance of a bird may be undetected in many species, yet strictly speaking the resultant plumage is alternate. Some birds (e.g. some grebes; Piersma 1988b) are in body-moult practically continuously. In large birds, body-moult is often prolonged or obscure. Then it can be impossible to identify plumages consisting of one generation of feathers.
Various terminologies have been used to categorize calendar age (Anon. 1985;Rogers 1989).None seems comprehensive enough for consistent application to all birds in a region extending from the Antarctic to the Tropics. The age of a bird in a given plumage is usually made clear in the moult sections; sometimes additional information is given next to the plumage headings.
Descriptions
Plumages are described in the sequence: head and neck, upperparts, tail, upperwing, underparts, underwing. Within these parts, we have generally worked from the front to the rear. In a few of the briefer descriptions, this sequence is modified to save space. Figures 1-21 illustrate the various parts. Some terms are defined in the glossary.
The appearance of individual feathers is described in the wing and tail. For other parts of the plumage, both feathers and general appearance of a given part are described. Where individual feathers are not described, the description of the tract also applies to the feathers (e.g. if a mantle is described as black, it consists entirely of black feathers). Terms used to describe patterns of feathers are shown in Figure 22.
The identity of colour depends on the light in which it has been seen and how precisely it needs to be defined. The greatest problem is that different people perceive colours differently. One man’s sepia is another’s fuscous, and so on. We have observed all skins in diffuse natural light or under a daylight globe (Phillips 60W Daylight). Names of colours given in the text are simple (e.g. dark brown) so that readers unfamiliar with complex colour descriptions (e.g. burnt umber with a trace of cinnamon) will not be misled. Where two colours are combined the last named colour is most dominant (e.g. buff-yellow is more yellow than buff); sometimes, the suffix ‘-ish’ is added to a colour, denoting a weaker tinge of that colour (e.g. buffish yellow has a weaker tinge of buff than buff-yellow). Further, light is darker than pale.
The F.B. Smithe Natwralist’s Color Guide (1975, 1981) has been used for descriptions of all colours. The identification numbers for the closest equivalent from this guide are bracketed in the text, after the simple names of colours. When the match is not particularly close the colour number is qualified, often with the abbreviation c. No numbers are given for colours which have no equivalent in the guide. If we have not been able to compare colours with the guide and if this is not clear from the context, we have denoted such colours (—). The names given to colours themselves in Smithe’s colour-guide have not been used; these were used by Smithe only to convey a sense of familiarity. Many (e.g. Pratt’s Payne’s Gray) are probably meaningless to most readers, and others ‘differ from widely held concepts’ (Pratt & O’Neill 1976).
From Vol. 2
Colour names given arc simple (e.g. dark brown) so that readers unfamiliar with more technical names (e.g. burnt umber) will not be misled. Wherever possible we have also used the FB. Smithe Naturalist’s Color Guide (1975, 1981) in describing colours. The identification numbers for the closest equivalent from this guide are bracketed in the text, after the simple names of colours, e.g. dark brown (121). When the match is not particularly close the colour number is qualified, often with the abbreviation ‘c’, e.g. dark brown (c 121). No numbers are given for colours which have no equivalent in the guide. If we have not been able to compare colours with the guide and this is not clear from the context, we have denoted such colours (— ). Where two colours are combined, the last-named colour is dominant (e.g. buff-yellow is more yellow than buff); sometimes the suffix ‘ish’ is added to a colour, denoting a weaker tinge of that colour (e.g. buffish yellow has a weaker tinge of buff than buff-yellow).
For some species, we have made mention of growth bars or fault bars as an ageing character. Growth bars are narrow bands of contrasting diffraction, thought to correspond to daily growth, which run roughly perpendicular to the shafts of most feathers. Like watermarks in paper, they are more conspicuous at some angles than others. They are more obvious in some feathers (especially rectrices) and some species than in others; the distance between bars and the width of each bar vary depending on rates of growth and other factors (for further information see Michener & Michener 1938; Murphy & King 199 1; Grubb 1992). If rectrices have characteristic growth bars or combinations of growth bars repeated identically on each feather, the feathers have grown simultaneously. This phenomenon of synchronic growth bars can also be seen in the secondaries but is most conspicuous in the tail (see Svensson [1984] for more information). Fault bars are narrower, translucent bands of similar orientation to growth bars, caused by defective formation of barbules; they occur only occasionally, perhaps as a result of a brief episode of stress or dietary deficiency (King & Murphy 1984). As with growth bars, fault bars will be synchronic only in tracts in which all feathers have grown simultaneously; both are thus most likely to be found in juvenile plumage. It should be noted that synchronic growth bars are of no use in ageing species (for instance, most Rallidae) which shed all their rectrices and remiges at one time in the course of pre-basic moult. In addition , synchronic tail bars will also develop when all rectrices arc replaced after accidental tail loss. Synchronic growth bars are thus usually supportive rather than conclusive indicators of juvenile plumage.
From Vol. 3
[paragraph on colour similar to previous volumes]
This Volume deals with many species that have complex sequences of moult, so we again present the nomenclature used for plumages and moults. All plumages and moults are given names corresponding roughly with the role they play in the life cycle, following the terminology used in BWP. The terms used will be familiar to most readers, but they can often be misleading, or imply relationships between specific plumages and moults with maturity or breeding, which have not been established or, in some cases, do not exist (see, for example, the introduction to Volume 1). The scheme of moult and plumage nomenclature developed by Humphrey & Parkes (1959, 1963; also summarized in the introduction to Volume 1) is more precise and usually less likely to be misleading. We have used both systems, which are summarized in Table 1, placing the Humphrey & Parkes nomenclature in brackets after the main heading. The conceptual differences between the two schemes are greater than the table below implies; see Humphrey & Parkes (1959), Thompson & Leu (1994) and references therein for further information.
From Vol. 5
[Paragraphs. 1, 3 and 4 repeated in Vol. 6 and Vol. 7, sometimes with minor rewording]
This section opens with a summary paragraph that states the Handbook editor, or editors who prepared this and subsequent sections and describes the overall sequence of plumages and moults from nestling to definitive plumage as far as it is known. The descriptions that follow are based on museum skins unless otherwise stated. Unlike earlier volumes, we now state the number of skins of each sex and age that were examined in preparing the descriptions, and the museum collections from which they came. Where more than one subspecies is described separately in Plumages, the number of skins are given separately for each subspecies. If other sources of information (e.g. examination of live birds) form a major portion of the descriptions, they are also noted here; otherwise they are simply given in the text.
Plumages are normally dealt with in the sequence: adult breeding, adult non-breeding, nestling or downy young, juvenile and immatures. When plumages differ between the sexes, males are generally described first, within each of these categories. Within each age and sex described, plumages are described in the sequence head and neck, upperparts, underparts, uppertail, undertail, upperwing, and underwing. Within these topographical areas, we try and work from the front to the rear. In a few of
the briefer descriptions, this sequence is modified to save space. Plumage areas are illustrated in the figures of topography. Some of the terms are defined in the glossary.
The appearance of tracts or topographical areas of the bird are described, with the appearance of individual feathers described as appropriate; individual feathers are frequently described for the wing and tail. Where individual feathers are not described, the description of the tract also applies to the feathers (e.g. if the mantle is described as black, it consists entirely of black feathers). Terms used to describe patterns of feathers are shown in the figures of topography.
The identity of colour perceived depends on the light in which it has been seen and how precisely it needs to be defined. The greatest problem is that different people perceive colours differently. We have observed all skins in diffuse natural light or under a daylight globe (Philips 60W Daylight). Colour names given are simple (e.g. dark brown) so that readers unfamiliar with more technical names (e.g. burnt umber) will not be misled. Wherever possible we have also used the EB. Smithe Naturalist’s Color Guide (Smithe 1975, 1981) in describing colours. The identification numbers for the closest equivalent from this guide are bracketed in the text, after the simple names of colours, e.g. dark brown (121) . When the match is not particularly close the colour number is qualified, often with the abbreviation ‘c’, e.g. dark brown (c121). For colours that have no equivalent in the guide, we place (ne) in brackets after the colour descriptor. If we have not been able to compare colours with the guide and this is not clear from the context, we have denoted such colours (—). Where two colours are combined, the last-named colour is dominant (e.g. buff-yellow is more yellow than buff); sometimes the suffix ‘ish ‘ is added to a colour to denote a weaker tinge of that colour (e.g. buffish yellow has a weaker tinge of buff than buff-yellow). We have not used the names given to colours themselves in Smithe’s colour-guide; these were used by Smithe only to convey a sense of familiarity. Many (e.g. ‘Pratt’s Payne’s Gray’ ) are meaningless to most readers, and others ‘differ from widely held concepts’ (Pratt & O’Neill 1976).
Growth-bars or fault-bars
For some species, we have made mention of growth-bars or fault-bars as an ageing character. GROWTH-BARS are narrow bands of contrasting diffraction that run roughly perpendicular to the shafts of most feathers and are thought to correspond to periods of daily growth. Like watermarks in paper, they are more conspicuous at some angles than others. They are more obvious in some feathers (especially rectrices) and some species than in others; the distance between bars and the width of each bar vary depending on rates of growth and other factors (for further information see Michener & Michener 1938; Murphy & King 1991; Grubb 1989,1991, 1992, 1995). If rectrices have characteristic growth-bars or combinations of growth-bars repeated identically on each feather, the feathers have grown simultaneously. This phenomenon of SYCHRONOUS GROWTH-BARS can also be seen in the secondaries but is most conspicuous in the tail (see Svensson [1984] for more information). FAULT -BARS are narrower, translucent bands of similar orientation to growth-bars, caused by defective format ion of barbules; they occur only occasionally, perhaps as a result of a brief episode of stress or dietary deficiency (King & Murphy 1984). As with growth-bars, fault-bars will be synchronous only in tracts in which all feathers have grown simultaneously; both are thus most likely to be found in juvenile plumage. It should be noted that synchronic growth-bars are of no use in ageing species (for instance, most Rallidae) that shed all their rectrices and remiges at one time in the course of pre-basic moult. In addition, synchronic tail-ars will also develop when all rectrices are replaced after accidental loss of the tail. Synchronic growth-bars are thus usually supportive rather than conclusive indicators of juvenile plumage.
BARE PARTS
[Information from Vol. 1, 2, 5]
From Vol. 1
These are described for the same subspecies as described in the plumages, in the sequence: iris; bare areas on head (if any); bill; leg and foot. Conventions for describing colours follow those used for plumages. Colours of bare parts cannot be studied satisfactorily in museum skins, because they generally change after death. We have therefore relied on colour photographs, our descriptions of recently dead, and live (mostly captive) birds, and on information on specimen labels or in literature.
From Vol. 2
Bare parts are now described in the sequence: bill, bare areas on head (if any), iris, leg and foot.
From Vol. 5
Bare parts are described in the sequence: bill, bare areas on head (if any), iris, and leg and foot.
MOULTS
[see Plumages for nomenclature discussion]
[Information from Vol. 1, 4, 5]
From Vol. 1
Nomenclature of moult was partly discussed above and summarized in Table 1. This section summarizes what is known of the pattern and timing of moult; where we have enough information, primary-moult is shown on the diagrams of annual cycle. Sequences of primary, secondary- and tail-moult are given where possible. Primary-moult from the carpal joint to the outside of the wing is called outwards; this sequence has often been called descendant in the past. Other terms describing patterns of moult are defined in the glossary. Conventions in numbering remiges and rectrices are described under Structure.
In species for which there are few data, we have recorded or scored moult of individual birds. Moult of remiges and rectrices is recorded with a system in which each feather is scored on a scale from 0 to 5 (Ashmole 1962; Ginn & Melville 1983). An unmoulted old feather is scored 0; a fully grown new feather is scored 5; a feather scored 1 is in pin; scored 2, less than one-third grown; scored 3, between one- and two-thirds grown; scored 4, two-thirds to fully grown. Primary-moult is recorded from the inside to the outside; superscripts denote the number of adjacent primaries of a particular score. For example, the shorthand N6311102 describes the primary-moult of a wing in which primaries 1 to 6 are new, primary 7 is between one- and two-thirds grown, primary 8 is in pin, and primaries 9 and 10 are yet to be moulted. Similar conventions are used for describing secondary- and tail-moults. Secondary-moult is recorded from the carpal joint inwards, and tail-moult from above, from the outer left to the outer right.
A moult-score is the sum of feather scores; we have sometimes applied it to primaries or the tail. In birds with 10 primaries the primary moult-score will lie between 0 (all feathers old) or 50 (all feathers new). Unless otherwise stated, we have scored the moult of both halves of the tail, and the primaries of one wing. We have not recorded condition of moult of the remicle, or used it in moult-scores.
From Vol. 4
MOULT-SEQUENCES
In species for which there are few data, or unusual patterns of primary-moult have been noted, we sometimes provide moult-sequences for individual birds. A widespread method of recording moult-sequences is to score each feather on a scale from 0 to 5 (Ashmole 1962; Ginn & Melville 1983) . Using this method, an unmoulted old feather is scored 0, and a fully grown new feather is scored 5; a feather in pin is scored 1; less than one-third grown, scored 2; between one- and two-thirds grown scored 3; and two-thirds to fully grown scored 4. Moult of primaries is recorded from the inside to the outside; in moult-sequences, the superscripts denote the number of adjacent primaries of a particular score, e.g. 54413104.
However, additional information, useful, for example, in ageing or determining the sequence of primary-moult, can be conveyed in moult-sequences by describing the state of abrasion of fully grown feathers. We have adopted a scoring system suggested by Rogers (1990) in which fully grown feathers are labelled with letters to indicate the state of wear of each feather: old feathers are labelled O; very worn feathers are labelled V; slightly worn feathers are labelled S; and new feathers are labelled N. Growing feathers are still scored from 1 to 4, as above. Thus, for example, a bird with a sequence of N24121O3V3 would have two new inner primaries, p3 and p4 are growing, p5 to p 7 are old and the outer three primaries are extremely worn. This bird has three different ages of fully grown primaries, and one interpretation of a sequence such as this is that the outer three primaries were not replaced in the last moult and are more than 12 months old. Similar conventions are used for describing moult of secondaries or tail. Moult of secondaries is recorded from the carpal joint inwards, and moult of the tail recorded from above, from the outer left to the outer right.
A primary moult-score (PMS) is a sum of the individual feather scores. In birds with ten primaries the PMS will lie between 0 (moult of primaries has not yet begun) or 50 (moult of primaries has recently finished) . When calculating PMS, feathers labelled O and V are scored 0; feathers labelled N are scored 5; and S can equal 5 or 0 depending on the circumstances. For example, a bird that has temporarily suspended moult of primaries may have a moult sequence of S4O6 in which case S = 5 and the PMS = 20. When this bird begins moulting again it may have the sequence S4N131O4, again S = 5 and the PMS = 28. However, a bird that has undergone a partial moult of outer primaries may have a moult sequence of O7S3. In this case, S = 0 and the PMS = O. Unless otherwise stated, we have scored the moult of the primaries of one wing. We have not recorded condition of moult of the remicle, or used it in moult-scores. [this para. repeated in Vol. 6 and Vol. 7]
REFERENCES
- Ashmole, N.P. 1962. Ibis 103B: 235-73.
- Ginn, H.B., & D.S. Melville. 1983. BTO Guide 19.
- Rogers, D.I. 1990. Corella 14: 141-7.
From Vol. 5
The nomenclature of plumages and moults has been discussed fully above. This section summarizes what is known of the pattern and timing of moult. Sequences of moult of primaries, secondaries, tail and body are given where possible. We have used the term OUTWARD to describe moult of
primaries from the carpal joint to the outside of the wing; this sequence has often been called descendant in the past; we have not used this confusing term because it is based on the out-dated practice of labelling the outermost primary the first (see discussion below in Structure). Other terms describing patterns of moult are defined in the glossary.
[ MOULT-SEQUENCES as for Vol. 4 above]
MEASUREMENTS
[For a complete list of measurement methods see Measurements Glossary]
[Information from Vol. 1, 2, 3, 4, 5, 7]
From Vol. 1
Measurements taken
For most species, measurements are given for length of wing, eight primary, tail, bill, tarsus and middle toe. Total length of head, and width and depth of bill have also been recorded for a few species. All measurements are recorded in millimetres. Our own measurements of dried museum skins are given. Measurements of skins and of live birds from the literature and other sources are presented separately, because there is variation in measurements taken by different individuals and because post-mortem shrinkage can have substantial effects on measurements. We used the methods described below. [For a complete list of measurement methods see Measurements Glossary]
Figures 1 to 22 – see Parts of birds
WING-LENGTH
Maximum chord of the wing was measured to the nearest millimetre: from the carpal joint to the tip of the longest primary. This section of the wing is flattened against a rule and straightened as much as possible; maximum chord is thus the longest measurement possible between the carpal joint and the tip of the longest primary. We did not measure wings in which the longest primaries had severe abrasion. Measurements of maximum chord can vary somewhat between measurers, especially because the experienced tend to take longer measurements (e.g. Jenni & Winkler 1989 and references therein).
Natural chord, and flattened chord, measurements of length of wing, are measured from the same points, but both give a considerably shorter reading. We have taken neither, but frequently quote from literature in which these methods have been used. In the former, no straightening or flattening of the wing is used (see Baldwin et al. 1931; BTO 1984); in the latter, the wing is only flattened (see Witherby et al. 1938; BTO 1984). Both were more widely used in the past, but are now believed to be less easy to repeat than maximum chord (e.g. Evans 1964; Ewins 1985), partly because primaries can straighten somewhat when wet (Evans 1964). Wing-length decreases in museum specimens. Shrinkage of between 0.39 and 3% has been reported (Herremans 1985). Most studies of shrinkage have been done on waders. In these, the amount of shrinkage is larger in species with longer wings (Engelmoer er al. 1983). There is a good deal of individual variation in amount of shrinkage. Thus, it is probably impossible to apply a single conversion factor to all species. Shrinkage continues in skins until they have dried out. Engelmoer er al. (1983) found this took over 2 years in some waders, but shrinkage may stop in as little as 2 months in some auks (Harris 1980; Ewins 1985). Slight increases in wing-length after drying out have been reported (Engelmoer et al. 1983). These are unexplained; possibly wings can be stretched, or ligaments cut, by frequent remeasuring. Our samples were not large enough for us to attempt to eliminate these sources of bias.
EIGHTH PRIMARY
This measurement was described recently (Berthold & Friedrich 1979). As far as we know, there are no publications on its use on A’asian birds. We took the measurement with a narrow ruler, inserted between the bases of the eighth and ninth primaries. The correct point of insertion can be seen by turning back the primary under wing-coverts. The distance from this point to the tip of the stretched eighth primary was recorded to the nearest millimetre. Jenni & Winkler (1989) recommend the use of a piece of folded graph-paper rather than a ruler; our measurements were taken before their paper was published.
Measurements of eighth primary are recommended by EURING, and they may supersede measurements of wing-length, being more exact and more easily repeated; there is considerably less variation between measurers; the measurement may be less likely to injure live birds (Berthold & Friedrich 1979; Jenni & Winkler 1989). In small European passerines, there is no post-mortem shrinkage in measurements of the eighth primary. It is not known if shrinkage occurs in larger birds. Piersma (19883) has shown that shrinkage in wing-length is not caused by a decrease in length of the feathers, but preliminary data of Jenni & Winkler (1989) suggest that some shrinkage occurs in large birds, presumably caused by contraction of skin between the primaries. Length of eighth primary and wing-length are closely correlated, and conversion factors from one to the other can be developed. In passerines, the eighth primary is measured rather than other primaries, because it usually has the closest correlation with wing-length. It is not known whether this is so in other groups but measurement of other primaries may be more appropriate in some groups. For reference throughout and for further information, see Jenni & Winkler (1989).
TAIL-LENGTH
Length of tail was measured to the nearest millimetre: as the distance between the point of emergence of the central tail-feathers from the skin to the tip of the longest feather. Post-mortem changes in tail-length are slight and not well understood; decreases (Greenwood 1979) and increases (Bjordal 1983; Herremans 1985) have been reported.
BILL-LENGTH
Length of bill was measured with calipers to 0.1 mm. We generally measured exposed culmen, the chord of the culmen from the bill tip to the frontal feathering. In some birds, the junction of the frontal feathering is not clearly defined, and other methods are used. BILL (S) is the chord of the culmen from the bill tip to the naso-frontal hinge. BILL (N) is the distance from the distal corner of the nostril to the bill tip. BILL (C) is the distance from the front edge of the cere to tip of bill. In procellariforms, the tip of the upper mandible is hooked so that in some the curve of the maxillary unguis, and not the tip, is farthest from the base of the bill. In these circumstances, culmen is measured to the curve of the unguis and not the tip.
Post-mortem changes in dimensions of the bill may depend on its structure (Fjeldså 1980); for example, no changes have been found in most waders studied (Greenwood 1979; Engelmoer et al. 1983), but significant decreases and increases as great as 2.9% have been reported in some species (Summers 1976; Engelmoer et al. 1983).
BILL-WIDTH, BILL-DEPTH
Depth of bill (BILL D) is measured from the junction of frontal feathering with the exposed culmen, to the lower edge of the mandibular ramus below; it is the minimum depth possible at this point. Width of bill (BILL W) is the distance between the tomia at the same point. Other conventions for taking these measurements are used for some species; these are given in the text. Width and depth of bill are subject to a great deal of shrinkage (Kinsky & Harper 1968; Fjeldså 1980) and are seldom used on skins.
TOTAL HEAD-LENGTH
Total head-length (THL) is measured to 0.1 mm with calipers, from the back of the skull to tip of bill. The measurement is becoming widely used for live birds because there is little variation between individual measurers. We have included it when data are available. THL cannot be taken consistently on skins, because the backs of their skulls are removed to differing extents during preparation.
TARSUS
Length of tarsus was measured to 0.1 mm from the midpoint of the hindside of the joint between the tibia and tarsus, to the midpoint of the joint between tarsus and middle toe in front. Shrinkage of tarsus is generally insignificant; Bjordal (1983) attributed the few reported post-mortem changes to difficulty in applying the measurement to exactly the same points in fresh and dried legs.
MID-TOE
Middle toe is measured from the joint at the base of the middle toe (in front of the leg) to tip of middle claw. In some groups it is customary to measure the middle toe without the claw. Length of toe decreases by about 2% after skinning (Fjeldså 1980; BWP [Vol. 5]). Its measurement is also difficult to standardize from museum skins because preparation and alignment of feet vary.
Presentation of Measurements
The tables of measurements are prefaced by information on the samples measured. Geographical location and age (where known) is recorded, and we mention whether measurements were taken on live birds or skins. The source of the data is given in brackets; all measurements of skins for which we simply give the institutes holding the specimens, were taken by R. O’Brien or D.I. Rogers. Museum sources are specified using our standard abbreviations (see Abbreviations and Conventions). Measurements of recently dead birds with similar citations were taken by the staff of the cited institutes. Except where stated, measurements were taken as above; if methods of measuring are unknown or different, this crucial fact is always mentioned. Different methods are described briefly. Some authors who published measurements without giving their methods have since described them to us. Measurements are presented in the standard fashion: mean (standard deviation; range; number of birds measured). Where sample sizes are of three or less, we present measurements of individuals. The statistical significance of the difference between two means is tested with a two-tailed t-test (e.g. Sokal St Rohlf 1969; Fowler & Cohen n.d.). When the probability of differences being due to chance (P) is less than 0.05, this is denoted *; when P<0.01, this is denoted **. Where values differ markedly from those given elsewhere, this is discussed.
From Vol. 2
Measurements of skins are given for nearly all species; details of the sources of data are given in brackets. All measurements of skins for which we simply give the institutes holding the specimens were taken by D.I. Rogers, D.J. James or A.M. Dunn. For species in which males and females look similar, sexing of skins has been based on data on labels. Several authors (e.g. Parkes 1963; Schodde et al. 1992) have pointed out that the sexes assigned on labels are not always accurate. In theory it should be possible to sex all fresh specimens by dissection; in practice it can be difficult, especially when birds are not in breeding condition. At such times their gonads are small, difficult to find and readily confused with the adrenal glands or other organs; when dissected by the inexperienced, mis-sexing can often occur. In addition, sexes recorded on labels of some old specimens (e.g. those collected by Robert Grant) appear not to have been based on dissection. We have reduced the numbers of mis-sexed birds in our samples by relying mainly on specimens for which gonads were sketched or described, and on specimens prepared by experienced workers, a process that can involve rather subjective decisions. A few mis-sexed specimens were probably not discarded on this basis; we have mentioned those few cases where it is felt that mis-sexed specimens may have affected the means calculated for either sex.
The following standard measurements have been taken for almost all species; the convention in reporting lengths of toe differs slightly from that in Volume . As in Volume 1, when quoting measurements from the literature we always mention cases where measuring methods differ from ours or are unknown. Differences between sample means were tested with two-tailed t-tests (if n >3). [For a complete list of measurement methods see Measurements Glossary]
WING
Length of the wing, measured to the nearest millimetre on the folded wing, from the carpal joint to the tip of the longest primary. We measure the maximum chord, flattening this section of wing against a butted ruler and straightening it as much as possible; maximum chord is thus the longest measurement possible between the carpal joint and the tip of the longest primary.
EIGHTH PRIMARY
Length of the eighth primary; measured with a narrow ruler inserted between the bases of the eighth and ninth primaries. The correct point of insertion can be seen by turning back the primary under wing-coverts. The distance from this point to the tip of p8 was recorded to the nearest millimetre.
TAIL
Length of tail, measured to the nearest millimetre with a ruler, as the distance between tie point of emergence of the central tail feathers from the skin to the tip of the longest tail feather.
BILL
Length of bill was measured with calipers to 0.1 mm; we generally measured exposed culmen, the chord of the culmen from the tip to the frontal feathering. In some groups the junction of the frontal feathering with the culmen is not clearly defined and a different method was used; these methods are given in the respective texts.
TARSUS
Length of tarsus was measured with calipers to 0.1 mm from the midpoint of the hindside of the tibio-tarsal joint, to the midpoint of the joint between tarsus and middle toe in front.
TOE C
Length of middle toe was measured with calipers to 0.1 mm from the joint at the base of the middle toe (in front of the leg) to the tip of the middle claw.
TOE
As TOE C, but excluding the middle claw.
REFERENCES
- Grubb, T.c. 1992. Auk 109: 673- 6.
- King, J.R., & ME Murphy. 1984 Auk 101: 168-9.
- Michener, H., & J.R. Michener. 1938. Condor 40: 149-60.
- Murphy, M.E. , & J.R. King. 1991. Auk 108: 695-704.
- Parkes, K.C. 1963. Living Bird 2: 121- 30.
- Schodde, R., et al. 1992. Corella 16: 23-8.
- Smithe, F.B. 1975. Naturalists Color Guide. Am. Mus. Nat. Hist., New
York. - — Naturalists Color Guide. Part III . Am. Mus. Nat. Hist., New York.
- Svensson, L. 1984. Identification Guide to European Passerines. Third Edition; Author, Stockholm (distributed by Br. Trust. Orn., Tring) .
From Vol. 3
[First and second paragraphs as for Vol 2 except for those taking the measurements] … were taken by K. Bartram, A.M. Dunn, D.J. James, D.I. Rogers, R.P. Scofield or A.J. van Loon.
[The following additional measurements were defined. See Vol 2 above for those repeated: WING, TAIL, TARSUS, TOE, TOE C. For a complete list of measurement methods see Measurements Glossary ]
BILL-LENGTH (Bill): measured with calipers to 0.1 mm; we usually measured exposed culmen, the chord of the culmen from the tip of the frontal feathering. In some groups the junction of the frontal feathering with the culmen is not clearly defined and a different method was used; these methods are given in the respective texts.
BILL-DEPTH (Bill D): measured from the junction of frontal feathering with the exposed culmen, to the lower edge of the mandibular ramus below; it is the minimum depth possible at this point.
BILL-WIDTH (Bill W): the distance between the tomia at the junction of frontal feathering with the exposed culmen.
Other conventions for taking measurements of depth and width of the bill are used for some species; these are given in the text. Width and depth of bill are subject to a great deal of shrinkage (Kinsky & Harper 1968; Fjeldså 1980) and are seldom used on skins.
TOTAL HEAD-LENGTH (THL): measured to 0.1 mm with calipers, from the back of the skull to tip of bill. The measurement is becoming widely used for live birds because there is little variation between individual measurers. We have included it when data are available. THL cannot be taken consistently on skins, because the backs of their skulls are removed to differing extents during preparation.
REFERENCES
- Fjeldså,J. 19S0. Bull. Br. Orn . Club 99: 143-5.
- Humphrey, P.S., & K.C Parkes. 1959. Auk 76: 1-31.
- —, — 1963. Auk SO: 496-503.
- Kinsky, F.C, & P.C Harper. 1965. Ibis 110: 100-2.
- Parkes, K.C 1963. Living Bird 2: 121-30.
- Rohwer, S., et al. 1992. Condor 94: 297-300.
- Schodde, R., et al. 1992. Corella 16: 23-S.
- Smithe, F.B. 1975. Naturalists Color Guide. Am. Mus. Nat. Hist., New York.
- — 1981. Naturalists Color Guide. III . Am. Mus. Nat. Hist., New York.
- Thompson, CW., & M. Leu. 1994. Condor 96: 769-S2.
- Willoughby, E.]. 1992. Condor 94: 295-7
From Vol. 4
Largely as in previous volumes. All measurements of skins made during this study (for which we simply give the institutes holding the specimens) were taken by K. Bartram, A.M. Dunn, D.J. James or D. I. Rogers. [rest of text repeated definitions of some measurements]
From Vol. 5
For most species, measurements are given for length of wing, tail, bill, tarsus and middle toe; total length of head, and width and depth of bill have also been recorded for some species. We no longer measure length of eighth primary (see Volume 1). Measurements of skins, taken by HANZAB researchers, are given for nearly all species. Published measurements from the literature or unpublished data of live birds or skins, are also given where they available, or references listed.
[text similar to text from previous volumes not repeated]
Presentation of measurements
[Text similar to Vol 1 except for those taking the measurements] All measurements of skins made during preparation of this volume (for which we simply give the institutes holding the specimens) were taken by K. Bartram, A.M. Dunn, J.S. Matthew, S.A. McKenzie or D.I. Rogers.
From Vol. 7
In addition to the standard measurements in HANZAB 5 we have used the following measurements in some species accounts:
BILL WN: Width of bill measured at anterior edge of nostrils
BILL DN: Depth of bill measured at anterior edge of nostrils
[For a complete list of measurement methods see Measurements Glossary]
WEIGHTS
Weights are taken from specimen labels, from published or unpublished data on birds captured for banding, and from other sources in the literature. Except where stated, weights are given in grams. They are presented in the same way as measurements. Information on variation in weight is summarized if available.
STRUCTURE
The following points are treated: shape of wing; number of primaries, secondaries, tertials and humerals (if present); wing formula (see Glossary); shape of tail, bill, and leg; other structural peculiarities. Primaries are numbered from the carpal joint outwards. Individual primaries are indicated by a ‘p’ and a number, p1 being the innermost, p10 the outermost functional primary in most species. The secondaries (including tertials, see glossary) are numbered from the carpal joint inwards, abbreviated as s1, s2 etc. The tail-feathers (retrices) are numbered from the central pair outwards: t1, t2 etc. All wing formulae were taken on skins; wing formulae can be affected by post-mortem shrinkage (Mead 1977; Knox 1980) but no other source of data was available.
AGEING, SEXING
Characters that can be used for ageing or sexing or both that have not been covered in Plumages or Bare Parts are described here [as are combinations of characters that distinguish ages or sexes. From Vol 5].
RECOGNITION
Sometimes included for species that are difficult to identify. This section deals with characters that are only usually visible in the hand; other characters are given in Field Identification.
GEOGRAPHICAL VARIATION
The general nature of the geographical variation is summarized (even where no forma subspecies are recognized) and differences between the recognized subspecies are given. An outline of taxonomic opinions on disputed treatments is usually given.
From Vol. 5
The general nature of the geographical variation is summarized (even where no formal subspecies are recognized) and differences between recognized subspecies, if any, are given, in reference to the detailed descriptions already given in the Plumages and Bare Parts sections. An outline of differing taxonomic opinions on treatments of taxa is usually given.
Leave a Reply
You must be logged in to post a comment.